В.В. Жерихин (1978) обратил внимание на любопытнейшую черту энтомофаун этого времени: многие семейства, характерные для мезозоя, обнаруживаются в них в последний раз, тогда как сколько-нибудь заметного появления новых групп не отмечается, и при этом появляется целый ряд представителей реликтовых для того времени (раннемезозойских и даже позднепалеозойских) групп. Это «всплывание реликтов» считают одним из наиболее ярких симптомов кризиса, охватившего среднемеловые сообщества: вымирание доминирующих раннемеловых групп (наиболее тесно связанных с существующей сукцессионной системой) создает экологический вакуум, который ненадолго заполняют древние формы, оттесненные к тому времени «на обочину жизни».
Складывающиеся на совершенно новой основе раннекайнофитные сообщества первоначально были несбалансированными и неустойчивыми. Слабость биоценотической регуляции и обилие незанятых экологических ниш вели к очень быстрой, взрывной, эволюции различных групп организмов (из насекомых здесь можно отметить высших бабочек и мух, паразитических перепончатокрылых, а также общественных насекомых – муравьев и термитов), по мере же насыщения биоценозов темпы эволюции снижались. Сложные трофические цепи в новых кайнофитных сообществах сформировались очень быстро (листья, поврежденные минирующими бабочками, сами эти бабочки и их высокоспециализированные паразиты из числа наездников Braconidae появляются в летописи почти одновременно, в сеномане) и в дальнейшем менялись мало. «По экологическому облику и систематическому составу , – заключает Жерихин (1980), – позднемеловые фауны значительно сильнее отличаются от раннемеловых, чем от кайнозойских, и могут рассматриваться как ранний, первоначально сильно обедненный, вариант этих последних. В сущности, по набору жизненных форм они имеют не так уж много отличий от современных».
Если вернуться к истокам среднемелового кризиса – самому возникновению покрытосеменных, – то здесь обнаруживается еще один интересный аспект, на который обратил внимание А.Г. Пономаренко (1997). Дело в том, что, по данным последних лет, серьезные эволюционные изменения среди насекомых начинаются несколько раньше широкого распространения покрытосеменных – с конца юры. Это, казалось бы, противоречит гипотезе о причинной связи между этими явлениями – но только на первый взгляд. Именно в это время начинается очередная «...зация» – ангиоспермизация голосеменных, и широко распространяются проангиоспермы (термин В.А. Красилова, 1989) – голосеменные с отдельными чертами цветковых (таковы, например, гнетовое Eoantha с крупными, явно энтомофильными «цветами», или беннетит Manlaia, имитирующий осоку – см. рисунок 52). «С точки зрения экосистемы» точное систематическое положение всех этих проангиоспермов не столь уж важно – все они могут в некоторой степени играть экологическую роль настоящих покрытосеменных.
Неудивительно, что параллельно с энтомофильными голосеменными появляются антофильные насекомые; имитаторам цветковых в растительном мире сопутствуют столь же многочисленные имитаторы бабочек среди насекомых (таковы, например, Calligrammatidae из сетчатокрылых, или Aneuretopsychidae из скорпионниц – см. рисунок 52). Важнее же всего то, что здесь складывается система с положительной обратной связью: наличие у некого беннетита комплекса опыляющих его насекомых стимулирует появление энтомофилии у его соседей-голосеменных, что, в свою очередь, расширяет экологическую нишу этих насекомых-опылителей, и т.д. Каждый шаг на этом пути увеличивает вероятность следующего и делает появление покрытосеменных (т.е. растений с полным синдромом ангиоспермных признаков) совершенно неизбежным; какая же конкретно группа голосеменных даст начало «настоящим цветковым» (равно как – какие именно из мекоптероидных насекомых породят «настоящих бабочек»), вообще-то говоря, не столь уж важно. Именно эти взаимно индуцируемые процессы Пономаренко и назвал «ангиоспермизацией мира».
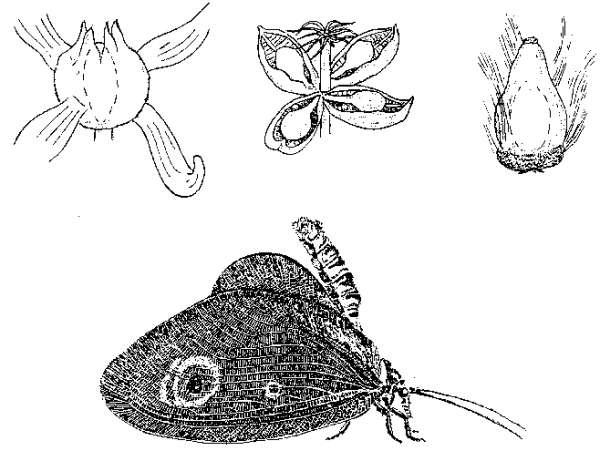
РИСУНОК 52. Голосеменные-имитаторы цветковых и насекомые-имитаторы бабочек. (а) – гнетовое Dinophyton, (б) – гнетовое Eoantha, (в) – беннетит Baisia, (г) – сетчатокрылое Kalligrammatidae.
«Ангиоспермизация мира» захватывает не только насекомых, но и все прочие компоненты континентальных экосистем. С начала мела на суше стремительно распространяются птицы (это особенно хорошо видно по укрупнению птерозавров – те полностью уходят в нишу крупных парителей) и териевые млекопитающие; в пресных водоемах настоящие костистые рыбы вытесняют ганоидов. После апт-альбского кризиса мир обретает вполне знакомый нам облик; правда, в нем наличествуют динозавры (и не просто «наличествуют» – они вполне процветает на протяжении большей части позднего мела), однако судьба этой группы была рассмотрена нами выше (глава 11), и нет оснований напрямую связывать ее вымирание с драматическими изменениями в морской биоте на границе мела и кайнозоя. Таким образом, мезозойско-кайнозойская смена на суше произошла много раньше, чем в море.
Ну, а каковы же всё-таки причины позднемелового «Великого вымирания»? Если не падение астероида и прочие «импакты» – то что же? Давайте прежде всего попытаемся найти какие-нибудь общие черты для этого события и для другого морского вымирания – пермско-триасового. Оно было самым крупным в истории Земли – тогда в морских группах вымерло 50% семейств, 70% родов и более 90% видов (для сравнения: при мел-кайнозойском вымирании исчезло менее четверти семейств). Помимо прямого исчезновения множества палеозойских групп (трилобиты, палеозойские кораллы – ругозы и табуляты) произошла радикальная смена структуры морских экосистем (роль основных рифостроителей перешла от мшанок к кораллам, донных фильтраторов – от брахиопод к двустворчатым моллюскам, и т.п.). Причины этой грандиозной фаунистической смены остаются загадкой; стандартные объяснения вроде «глобального катастрофического вулканизма» или морской регрессии, резко сократившей площадь шельфов (основной зоны обитания морских организмов) производят впечатление явно придуманных «за неимением лучшего». Мне лично наиболее остроумной и логичной (хотя и не бесспорной) представляется гипотеза Х. Таппан – специалиста по фитопланктону.