Если задать ботанику вопрос – какие черты покрытосеменных привели их к эволюционному успеху, он наверняка начнет говорить о двойном оплодотворении, о триплоидном эндосперме и т.п.; это всё, конечно, верно – но немножко не о том. «С точки зрения биосферы» наиболее существенны, пожалуй, три свойства цветковых, позволившие им радикально изменить самую структуру мезозойских экосистем.
1/ Склонность к неотении (неотения – обретение организмом половозрелости на ранних (личиночных) стадиях своего индивидуального развития), то есть – в данном конкретном случае – к образованию травянистых форм, которые по сути дела есть древесные проростки, ставшие способными к размножению; напомним, что голосеменные «траву» не образуют вовсе – ни сейчас, ни в прошлом. Именно травянистые двудольные являются самыми агрессивными эксплерентами («сорными растениями»), мгновенно захватывающими места нарушений растительного покрова, а травянистые однодольные, склонные к образованию многолетней дернины, наиболее эффективно препятствуют эрозии.
2. Появление «мясистых» листьев с большим объемом фотосинтезирующей паренхимы, вызвавшее появление листогрызущих насекомых. Дело в том, что до того насекомые потребляли лишь генеративные органы, содержимое сосудов, даже древесину – но только не листья: в листе голосеменного (даже такого, как гинкго), попросту говоря, нечего есть. Ситуация менятся лишь с появлением цветковых – объеденные листья и мины (ходы, проедаемые личинками насекомых в толще листа) известны с самого начала позднего мела (сеноманский век). На покрытосеменных сформировалась общирная фауна листогрызущих насекомых, и уже после этого они перешли на голосеменные, папоротники и хвощи.
3. Энтомофилия цветков, открывающая покрытосеменным путь к сложной коэволюции с насекомыми. Собственно говоря, сам цветок покрытосеменных, судя по всему, сформировался в процессе адаптаций к насекомоопылению для защиты завязи от повреждений ее опылителями.
Ангиоспермы впервые появляются в палеонтологической летописи в начале раннего мела, в неокоме; на протяжении барремского и аптского веков они представлены единичными находками, в альбе идет стремительное нарастание их числа, а уже в самом начале позднего мела, в сеномане, цветковые составляют большую часть ископаемых флор – то есть наступает «кайнофит» (рисунок 50). Среднемеловая экспансия цветковых была столь стремительной (она фактически уложилась в ничтожный по геологический меркам отрезок от 115 до 108 млн лет), что Дарвин в свое время в сердцах назвал этот эпизод палеонтологической истории «отвратительной тайной – abominable mistery». Делались попытки объяснить это тем, что цветковые на самом-то деле появились на Земле давным-давно, минимум в юре – просто они обитали лишь в горах (не попадая по этой причине в палеонтологическую летопись), а в середине мела заселили, наконец, приводные местообитания, благоприятствующие попаданию в захоронения. Вообще такой «период скрытого существования» сбрасывать со счетов никак нельзя – но в данном случае эта гипотеза не проходит: дело в том, что пыльца покрытосеменных (которая, ясное дело, летает везде, и ее не «упрячешь» в горы) появляется в летописи одновременно с их макроостатками. По всем признакам, цветковые действительно возникли в начале мела, причем именно в приводных местообитаниях.
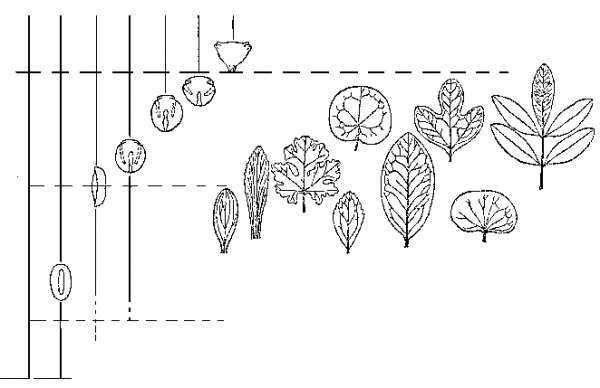
РИСУНОК 50. Распределение в меловых отложениях основных типов пыльцы и листьев покрытосеменных.
В 70-х годах советские палеоэнтомологи исследовали множество местонахождений меловых насекомых в Сибири и Монголии, «закрасив» тем самым самое крупное «белое пятно» в геологической истории класса. Оказалось, что у насекомых график, который отражает долю вымерших таксонов в фаунах разных времен, ведет себя довольно неожиданным образом (рисунок 51). Доля вымерших семейств мало меняется на протяжении большей части мезозоя, и на границе юры и мела составляет около 60% ; затем, в течении раннего мела, доля эта стремительно падает от 60 до 20% , в позднем мелу это падение резко замедляется (от 20 до 5%), а к началу кайнозоя кривая вновь «выходит на плато», только уже в районе 5%. Иными словами, у насекомых самая резкая фаунистическая смена, связанная с массовым вымиранием мезозойских групп, приходится не на конец мела, а на его середину – в точности как у растений. На этом основании В.В. Жерихин (1978) предложил следующую модель среднемелового биоценотического кризиса.
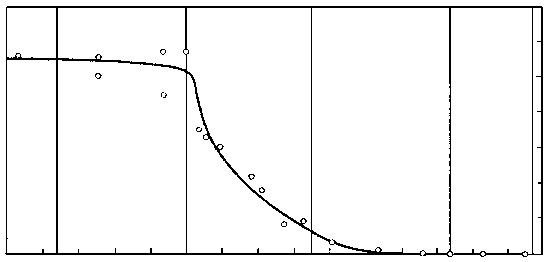
РИСУНОК 51. Эволюция насекомых в мезозое: процент вымерших семейств от общего числа семейств насекомых в позднемезозойских и кайнозойских фаунах.
Покрытосеменные возникли в начале мелового периода, первоначально как ценофобы – растения, не входящие в закономерные, сложившиеся в длительной коэволюции сукцессионные ряды[69] . Они росли в качестве «сорной» растительности по свободным от других растений участкам (на речных отмелях, береговых оползнях, гарях), которые соседствовали с гораздо более обширными участками, занятыми зрелыми сообществами мезофитной растительности. Покрытосеменные были исходно энтомофильными и смогли сформироваться лишь в окружении энтомофильных же голосеменных (таких, как беннетиты, саговники, кейтониевые), используя уже сложившийся к тому времени комплекс их опылителей. Но если покрытосеменные оказались среди пионерных растений мезофита единственными энтомофилами (а, судя по всему, так оно и было), то это должно было дать им гиганские преимущества: именно для локально распределенных, не образующих сплошных зарослей пионеров насекомоопыление особенно выгодно; впрочем, возможно, имело место и прямое конкурентное вытеснение покрытосеменными энтомофильных голосеменных из пионерных сообществ – за счет более эффективного привлечения насекомых, лучшей защиты от повреждения завязи опылителями и т.д.
Основой стремительной среднемеловой экспансии цветковых стало то, что им удалось закрепиться в качестве нормальной пионерной растительности – для чего решающими факторами стали их исходная энтомофильность и наличие среди них травянистых форм (что во много раз убыстряет зарастание поврежденных участков). Покрытосеменные не пытались потеснить всю мезофитную растительность, что было абсолютно нереально; они «всего-навсего» конкурентно вытеснили прежних пионеров – и тем самым блокировали все последующие стадии мезофитной экогенетической сукцессии. Разрушение существовавших в то время закономерных сукцессионных рядов (попросту говоря – мезофитная растительность продолжала существовать там, где она существует, но потеряла способность восстанавливаться после экзогенных нарушений), вызвало полный развал мезозойских наземных экосистем и массовое вымирание входивших в них животных; наиболее интенсивно эта деструкция шла в альбе (захватывая конец апта и начало сеномана).