6. Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент.
Прежде чем непосредственно приступать к изучению древнейших многоклеточных организмов, давайте задумаемся: а зачем, собственно говоря, эта многоклеточность нужна? Какие преимущества она дает? На эту проблему можно посмотреть с разных точек зрения, но мы для начала посмотрим – с экологической. Иными словами: что многоклеточность дает не самому организму, а экосистеме, и – в конечном счете – биосфере.
Малоразмерные фитопланктонные организмы (прежде всего – прокариотные) сталкиваются с одной проблемой, на которую впервые обратил внимание гидробиолог Б.В. Виленкин. Снабжение этих одноклеточных биогенами и растворенными газами происходит за счет диффузии сквозь клеточную стенку; это, казалось бы, должно благоприятствовать миниатюризации клеток – чтобы максимально увеличить отношение ее поверхности к объему. Однако очень мелкий организм неспособен выделиться из окружающей его водной массы: пассивно паря в толще воды (и оставаясь неподвижным относительно нее), он быстро создает вокруг себя «пустыню» – выедает из непосредственно окружающего его водного слоя все биогены. При этом он насыщает воду диффундирующей наружу органикой, которую не в силах удерживать внутри клетки из-за того же самого – высокого – соотношения поверхность/объем. То, что при этом безвозвратно теряется более трети клеточной продукции – полбеды; хуже то, что на окисление этой органики расходуется кислород, и водный слой, окружающий организм, эвтрофицируется. Одним словом, незабвенный Пятачок был прав: жизнь «очень маленького существа» полна неприятностей.
Неприятностей этих можно избежать, лишь заставив организм перемещаться относительно вмещающей его воды. Этого можно добиться двумя способами: либо начать самому активно двигаться в ее толще, либо, наоборот, прикрепиться к неподвижному субстрату – чтобы вода двигалась относительно тебя. Второй способ прокариоты реализовали немедленно, сконструировав мат. А вот с первым способом (казалось бы, более простым и очевидным) у них возникли крупные проблемы – ибо эффективных органов движения, подобных жгутикам и ресничкам эукариотных одноклеточных, у прокариот не возникает, а создание клеточных агрегаций, способных к согласованным движениям (например, волнообразным) затруднено из-за крайней слабости межклеточных взаимодействий (см. Главу 5). Поэтому «генеральной линией» этот способ становится лишь при появлении эукариот, способных к образованию высокоинтегрированных клеточных агрегаций, т.е. к настоящей многоклеточности.
Многоклеточность привела, среди прочего, к резкому повышению способности организмов создавать в своем теле запас питательных веществ. Практически лишенные этих запасов одноклеточные (в особенности – прокариоты) обречены реагировать на изменения содержания биогенов в окружающей среде единственным способом – изменением плотности популяций. (Примером такого их поведения может служить «цветение» водоемов, когда стремительное размножение одноклеточных и нитчатых водорослей быстро приводит к исчерпанию ресурса экосистемы; в результате безмерно разросшаяся популяция в одночасье вымирает, а на окисление этой мертвой органики расходуется затем почти весь кислород водоема – «замор»). Запасные вещества и резервная биомасса крупных организмов делают их популяции более независимыми от колебаний ресурса и стабилизируют их плотность. В свою очередь, существующие в экосистеме геохимические круговороты обретают в лице этих организмов крупное резервное депо[14] , оказывающее на все эти циклы мощное стабилизирующее воздействие. Такая «взаимная стабилизация» кажется одним из главных экологических следствий возникновения многоклеточных (хотя правильнее сказать – макроскопических, т.е. видимых невооруженным глазом) организмов.
Последняя оговорка не случайна. Если же мы от вопроса «для чего возникла многоклеточность» перейдем к вопросу «как и когда она возникла», то нам прежде всего придется определить – какой организм следует считать многоклеточным? Если даже не брать в расчет чисто терминологические проблемы (следует ли называть «колонией» Volvox, у которого есть дифференциация между клетками переднего и заднего полушарий, зародыша, возникающие путем палинтомии – деления без последующего увеличения размеров клеток, и который к тому же смертен), здесь существует и проблема вполне объективная. А именно: соответствует ли строгим критериям многоклеточности строение тела макроскопических водорослей и грибов? Большинство биологов ныне отвечают на этот вопрос отрицательно, оставляя термин «многоклеточные» лишь за зелеными растениями (Metaphyta) и животными (Metazoa); Дж.Корлисс (1983) ввел для этих двух групп удачный термин – многотканевые организмы.
Таким образом, достижение живыми организмами макроскопических размеров может происходить различными способами, а истиная многоклеточность (многотканевость) – лишь один из них. В любом случае, многоклеточность (в широком смысле) почти наверняка возникала в процессе эволюции многократно и независимо: во многих типах водорослей – красных (Rhodophyta), зеленых (Chlorophyta), золотистых (Chrysophyta) – или высших грибов-аскомицетов (Ascomyceta) можно выявить преемственные ряды от одноклеточных форм через колонии (например, нитчатые формы) к макроскопическим организмам с аналогами тканевой дифференцировки.
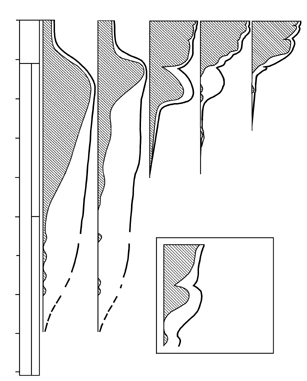
РИСУНОК 19. Изменения во времени относительного обилия основных биотических компонентов: (а) – прокариоты-строматолитообразователи, (б) – свободноживущие прокариоты, (в) – эукариотный фитопланктон, (г) – многоклеточные растения, (д) – многоклеточные животные.
Эта гипотетическая эволюционная картина хорошо соответствует современным палеонтологическим даным (рисунок 19). Первые эукариоты появились (как мы помним из главы 5) около двух миллиардов лет назад среди фитопланктонных акритарх; вскоре к ним добавились и нитчатые формы с эукариотными параметрами клеток. Замечательно, что они никогда не встречаются в прокариотных бентосных сообществах (цианобактериальных матах), а с самого начала формируют свой собственный тип растительности, названный В.Шенборном (1987) «водорослевыми лугами». Ныне сообщества такого типа известны лишь в некоторых антарктических внутренних водоемах; в докембрии же они, судя по характеру осадков, были широко распространены в морях за пределами мелководий (которые были заняты матами).