расслабление ретракторов, втягивавших ногу и щупальца; активация секреции слизи, необходимой для ползания по субстрату; учащение сердцебиений и т. д. Важнейший и остававшийся до сих пор незамеченным результат исследования клеточных механизмов заключается в том, что все компоненты этого поведенческого ответа обязаны посредничеству одного и того же медиаторного вещества – серотонина (5-НТ). При этом 5-НТ, секретируемый специфическими нейронами, управляет своими клеточными мишенями как контактно, так и дистантно: эффект 5-НТ на нейронный генератор жевательной моторики описывается в понятиях синаптической передачи, но при действии на некоторые другие мишени 5-НТ выступает в роли нейрогормона.
Из анализа клеточных механизмов следует, что 5-НТ является здесь фактором, интегрирующим поведение. Эта интегративная функция не обязательно осуществляется на каком-то определенном уровне иерархической структуры нервной системы моллюска: она реализуется и на уровне нейроэффекторной передачи (как, например, в случае управления сердцебиениями), и на уровне командного нейрона (управление моторикой радулы), и т. д. Существенно, что отдельные системы, участвующие в выполнении рассматриваемого поведенческого акта, в том или ином звене обеспечены такими рецепторами 5-НТ, которые позволяют этим системам отвечать на 5-НТ синергично. Разнообразие клеточных рецепторов 5-НТ и специфическое распределение этих рецепторов служат, таким образом, тому, чтобы разные клетки и органы участвовали в целостном поведении согласованно.
По нашей интерпретации, адекватные стимулы, вызывающие «реакцию пробуждения», посредством сенсорных клеток активируют 5-НТергические нейроны, вслед за чем 5-НТ становится фактором, определяющим поведенческий ответ на раздражение. В эксперименте активацию 5-НТергических нейронов можно имитировать введением 5-НТ в полость тела улитки: экзогенный 5-НТ вызывает хорошо скоординированное поведение, подобное «реакции пробуждения».
Интеграцию поведения улитки серотонином мы склонны рассматривать как пример широко распространенного класса явлений. Имеются разные основания считать, что другие продукты нейронной секреции принимают аналогичное участие в механизме управления поведением в простых нервных системах беспозвоночных. Известная амбивалентность всех медиаторных веществ становится понятной в контексте рассматриваемого механизма интеграции: нейротрансмиттер, оказывающий только возбуждающее или только тормозящее действие на разные мишени, не был бы способен выполнять роль интегрирующего фактора.
Идеализируя, поведенческий репертуар можно представить как набор синергических паттернов, каждый из которых управляется специфическим медиаторным веществом (точнее, продуктом нейронной секреции, способным действовать и контактно и дистантно). Такое представление о функционировании идеальной нейронной системы можно выразить формулой: один медиатор – одна синергия. Тогда гетерогенность нейронной системы становится мерой разнообразия поведения.
Сказанное касается простых нервных систем беспозвоночных, но сходным образом можно оценивать значение клеточной гетерогенности для сложных нейронных образований, таких как мозг млекопитающего, где те же рассуждения приложимы к локальным блокам нейронов. Идеализированная таким способом локальная нейронная система (например, модуль коры головного мозга) функционирует паттернами активности, при этом разнообразие паттернов (т. е. возможных состояний системы) определяется химической гетерогенностью источников секреции (т. е. афферентных входов и собственных интернейронов).
Предлагаемая идеализация позволяет концептуализировать явление химического разнообразия нейронов и синапсов, на этой основе могут строиться теоретические модели гетерогенных нейронных систем.
1985
Бессинаптическая идеализация нейронного ансамбля
[Доклад на конференции «Простые нервные системы»]
Сахаров Д. А. Синаптическая и бессинаптическая модели нейронной системы // Простые нервные системы. Ч. 2. Казань: КГУ, 1985. С. 78–80
Вкаждом локальном участке нервной ткани осуществляются быстрые, контактные взаимодействия между нейронами, традиционно описываемые в понятиях химического синапса. Изучение нейробиологической реальности на материале простых нервных систем позволяет переосмыслить основной постулат синаптической концепции – представление о синапсе как канале связи между нейронами. Концепцию анатомических (т. е. синаптических) каналов связи предлагается рассматривать как идеализацию, т. е. предельный, а не общий случай; в качестве противоположного теоретического предела выдвигается представление об идеальной бессинаптической системе, в которой избирательность контактных взаимодействий обеспечивается гетерохимичностью системы и химическими каналами связи. (Под идеализацией здесь подразумевается известный прием анализа, когда рассматривают теоретическую модель объекта, пренебрегая какими-то из его реальных свойств.)
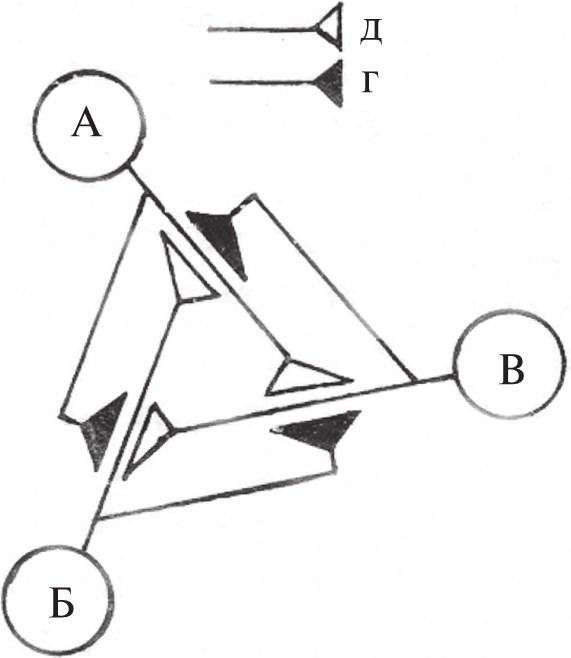
Рассмотрим обе идеализации на примере системы из трех нейронов – А, Б и В, связанных таким образом, что система функционирует как трехфазный генератор (рис. 1). Нейрофизиологическая феноменология такого рода характерна для реальных нейронных систем (в частности, для нейрональных генераторов ритмической мышечной активности). Интерпретируя, как это принято, такие системы в понятиях синапса, им приписывают свойство анатомической упорядоченности, связывая так наз. постсинаптические потенциалы и упорядоченную нейронную активность с наличием специфических контактов, как это сделано на рис. 2. По синаптической модели, сферой действия медиатора является синаптическая щель – компартмент межклеточного пространства, входящий в состав анатомического канала связи, синапса. Эта особенность модели позволяет пренебрегать разнообразием медиаторов: для функционирования рассматриваемой (рис. 2) и любой другой, сколь угодно сложной, нейронной системы необходим и достаточен один медиатор. В самом деле, при описании нейронных систем знаниями о химизме нейронов принято пренебрегать как несущественными.
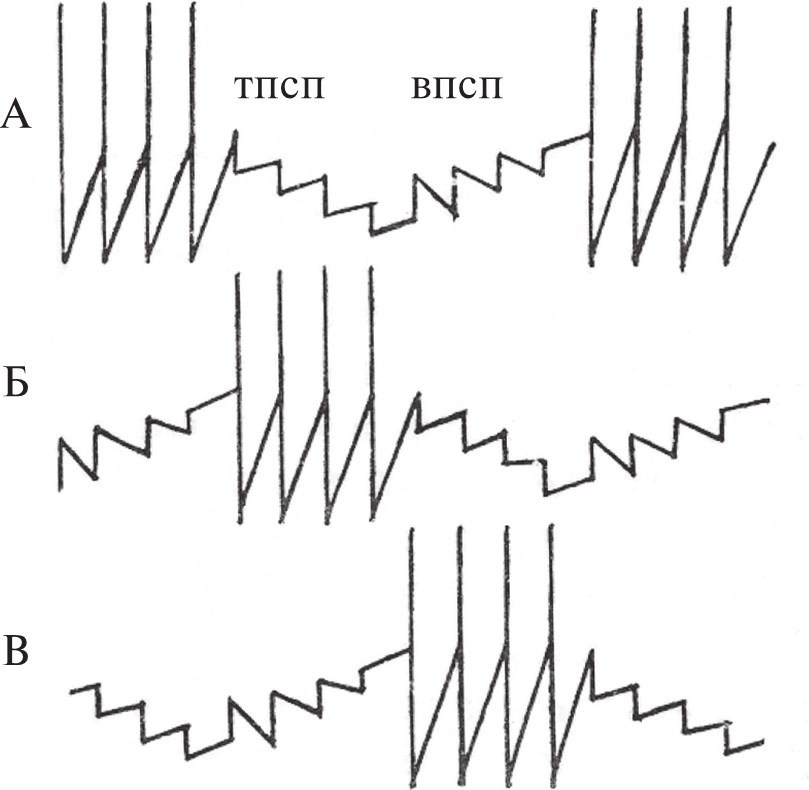
Рис. 1
Рис. 2.
где д – деполяризующее окончание, г – гиперполяризующее окончание
Предположим теперь, что система лишена специфических контактов:
вся поверхность каждого из трех нейронов наделена одинаковыми секреторными и рецепторными свойствами, отростки контактируют случайным образом, переплетаясь в общем, не разделенном на компартменты, внеклеточном жидком матриксе. Сможет ли такая бессинаптическая система по-прежнему функционировать так, как показано на рис. 1? Сможет – при условии, что каждый из трех нейронов выделяет собственный медиатор и имеет нужный тип рецепторов к медиаторам двух других нейронов.
Если медиаторы клеток А, Б и В обозначим как а, б и в; рецепторы к а, обеспечивающие де- и гиперполяризацию, как соответственно Рда и Рга и т. п., то эту гетерохимическую нейронную систему можно записать следующим образом:
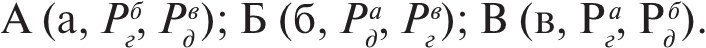
Такая запись содержит столь же полные сведения о специфических связях, обеспечивающих фазировку активности нейронов рассматриваемой системы, как и рис. 2, но содержание этих двух записей существенно различно. Бессинаптическая модель, в отличие от синаптической, составлена из качественно разнородных клеточных элементов (и в этом смысле подобна реальным нейронным системам, которые всегда гетерогенны); это усложнение сопряжено с такими преимуществами, как простота конструкции (неструктурированный, анатомически не упорядоченный нейропиль, плексус и т. п.) и простота управления (медиатор выполняет роль фактора, интегрирующего систему в целом). Интегративную функцию медиаторных веществ, присущую бессинаптической модели, трудно продемонстрировать на высших животных, в мозге которых эта функция может быть разной в разных участках нервной ткани. Вместе с тем у просто организованных животных эта функция может иметь специфическое выражение на поведенческом уровне. Действительно, интеграция поведения медиаторными веществами показана нами и другими авторами для ряда беспозвоночных. Эти данные указывают на близость реальных нервных систем (по крайней мере, у беспозвоночных) к бессинаптической модели.