Однако искусственные нейронные сети и структурная коннектомика всё еще имеют очень мало общего с процессами, происходящими в живых нервных системах. Эти модели основываются на идее «проволочного мозга», в котором он представляется электрической сетью с жестко заданной топологией, которая образуется проводами, соединяющими одинаковые «пассивные» нейроны. Многие важные свойства, присущие живым нейронным сетям, в таких терминах описать невозможно. Современные исследования свидетельствуют о том, что (1) нейроны не одинаковы – они являются трансмиттер-специфическими; (2) большинство нейронов способны генерировать собственную электрическую активность, а не только в ответ на возбуждающий внешний сигнал; (3) выброс трансмиттеров происходит в общее межклеточное пространство, а не только в синаптическую щель; (4) перестройка топологии сетей и изменение режимов активности нейронов происходят ad hoc – под действием разных нейротрансмиттеров [1, 4, 5].
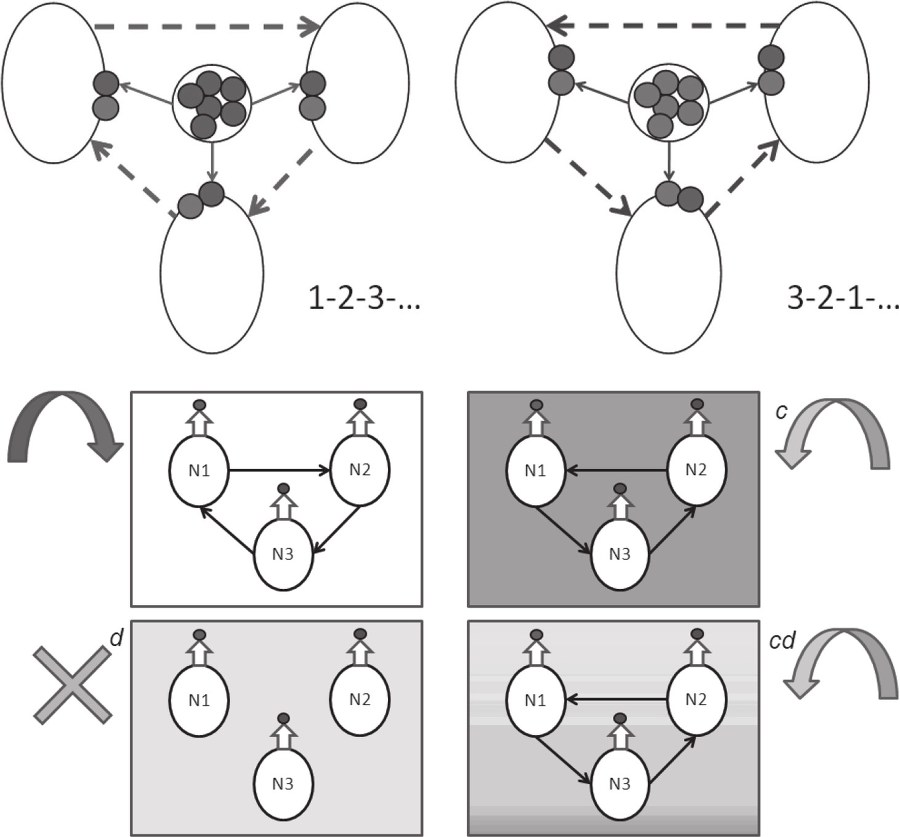
Л. Жилякова (ИПУ РАН) предложила математическую модель, которая развивает концептуальную модель гетерохимического генератора, предложенную еще тридцать лет назад Д. А. Сахаровым [3]. Модель учитывает принципиальные для функционирования реальной системы свойства биологических нейронов. Ее формальный нейрон обладает не только электрическим зарядом, но и химическими входами (рецепторами к разным нейротрансмиттерам) и химическим выходом (секретируемый нейротрансмиттер). Нейроны в этой модели «разноцветны», отличаются по входам и выходам, т. е. секретируют разные нейротрансмиттеры и по-разному реагируют на разные химические сигналы. Кроме того, они «не пассивны», обладают собственной, а не только вызванной активностью. Сеть из таких нейронов названа гетерогенной нейронной сетью. При этом термин «сеть» не означает наличия лишь электрических связей – любая химическая связь в ней может быть отражена теми же математическими средствами. Каждый значимый в этой сети трансмиттер может выделить свою подсеть. Переключение между сетями производится с учетом пороговых характеристик. Модель, обладающая такими свойствами, порождает сети с топологией, способной динамически изменяться при изменении концентраций тех или иных трансмиттеров в среде.
Уже сформулирована модель для простого моторного генератора (3 нейрона, 3 фазы ритма, 2 внутренних и 2 внешних нейротрансмиттера, способность к перестроению). Эта модель соответствует реальным биологическим характеристикам известных центральных генераторов паттерна (Central Pattern Generators). В частности, появление «эндогенной активности» нейронов автоматически привело к доминированию тормозных (как в биологических генераторах), а не возбуждающих (как в прежних сетевых моделях) связей. Трансмиттер-специфические перестройки сходны с наблюдаемыми в реальности благодаря наличию межклеточной среды и разноцветного нейромодуляторного фона. И наконец, жесткие синаптические связи оказались не необходимыми для работы модели, такой генератор «связан» химически изначально, что соответствует реальному онтогенезу нервной системы, в которой нейротрансмиттерная специфичность задается до установления синаптических связей. Помимо перечисленных преимуществ, модель дала ответ на частный нейробиологический вопрос: как трехфазный генератор может работать не на трех, а на двух нейротрансмиттерах?
Основная работа – еще впереди. На сегодняшний день важно то, что шаг в этом необходимом и правильном направлении наконец сделан. Нейробиологи и медики могут надеяться на биологически адекватное математическое моделирование реальных нейронных взаимодействий. А поскольку уже есть первые попытки создания физических аналогов хемочувствующих и хемо-секретирующих нейронов для протезирования нервной системы, появляется вероятность использования гетерогенной нейронной сети не только в фундаментальной, но и в прикладной науке.
Литература
[1] Дьяконова В. Е. Нейротрансмиттерные механизмы контекст-зависимого поведения // Журн. высш. нервн. деят. 2012. Т. 62. № 6. С. 1–17.
[2] Жилякова Л. Ю. Сетевая модель распространения нескольких видов активности в среде сложных агентов и ее приложения // Онтология проектирования. 2015. Т. 5. № 3 (17). С. 278–296.
[3] Сахаров Д. А. Синаптическая и бессинаптическая модели нейронной системы // Простые нервные системы. Ч. 2. Казань: КГУ, 1985.С. 78–80.
[4] Сахаров Д. А. Биологический субстрат генерации поведенческих актов // Журн. общ. биол. 2012. Т. 73. № 5. С. 334–348.
[5] Bargmann C. I. Beyond the connectome: how neuromodulators shape neural circuits // Bio-essays. 2012. No. 34. P. 458–465.
[6] Baronchelli A., Ferrer-i-Cancho R., Pastor-Satorras R., Chater N., Christiansen M. H. Networks in Cognitive Science // Trends in Cognitive Sciences. 2013. Vol. 17. No. 7.
[7] Bullmore E., Sporns O. Complex brain networks: graph theoretical analysis of structural and functional systems // Nature Reviews Neuroscience. 2009. Vol. 10. P. 186–198.
[8] McCulloch W. S., Pitts W. A logical calculus of the ideas immanent in nervous activity. Bull. Math. Biophys. 1943. Vol. 5. P. 115–133. (Рус. пер.: Маккаллок У. С., Питтс У. Логическое исчисление идей, относящихся к нервной активности // Автоматы / Под ред. К. Э. Шеннона и Дж. Маккарти. М.: ИЛ, 1956. С. 362–384.)
2017
Феномен ниши
[Доклад на конференции «Когнитивная наука в Москве: новые исследования»]
Сахаров Д. А., Дьяконова В. Е. и Жилякова Л. Ю. Ниша – механизм управления альтернативным выбором: от стволовой клетки к ансамблю нейронов // Когнитивная наука в Москве: новые исследования. М.: Буки-Веди, 2017. С. 322–326
Казалось бы, биология развития не имеет отношения к проблемам когнитивистики. Тем не менее в качестве завязки этого сообщения хотим привлечь внимание участников конференции к механизму клеточной дифференцировки. Находясь в точке бифуркации, стволовая клетка сама делает выбор между доступными ей путями специализации, например решает, стать ли ей нейроном или клеткой глии. Главную роль в механизме принятия решения играет микроокружение стволовой клетки – ее ниша. Это она направляет клетку по пути наименьшего сопротивления к одному из устойчивых состояний. Для тех, кто занимается стволовыми клетками, ниша не новость (см. обзоры [11], [10] ). Один из специалистов определяет нишу так: она – «a basic unit of tissue physiology, integrating signals that mediate the balanced response of stem cells to the needs of organisms» [14]. При небольшом усилии воображения мы обнаружим, что такое определение идеально приложимо к среде, омывающей паттерн-генерирующий нейронный ансамбль.
В этом нет ничего неожиданного. Ансамбли, собираемые из живых нервных клеток, имеют больше общего как раз со стволовыми клетками, чем с формальными или искусственными нейронами. Потому вполне естественно, что выбором из доступного ансамблю репертуара стабильных конфигураций управляет ниша. Наглядный пример – выбор локомоторного аллюра. В нашем коллективе было показано, что у морского ангела выбор из репертуара аллюров определяется балансом дофамина и серотонина в среде, омывающей генератор локомоции [6, 9]. Накопилось немало результатов, свидетельствующих о том, что и у других модельных объектов нейроэтологии нейротрансмиттерный состав среды, будучи транслятором поведенческого контекста, управляет выбором между альтернативными моторными программами (см. обзоры [13, 3] ).
Можно ли извлечь из знаний о нише выводы, значимые для понимания ментальных актов? Согласно правдоподобной гипотезе Энн Грейбил [7], «генераторы когнитивных паттернов» подобны тем нейронным ансамблям, какими продуцируются локомоторные и иные моторные акты, – хорошо изученным central pattern generators (CPGs) [5, 2].