1.1. Основные постулаты «теории»
I. Отбор половой есть нечто иное, чем отбор естественный. Вот что по этому поводу писал сам Дарвин: «Если оба пола ведут совершенно одинаковый образ жизни, и все-таки самец обладает органами чувств и перемещения, выше развитыми, чему самки[18], то может случиться, что усовершенствование этих органов необходимо самцу для отыскивания самки; но в огромном большинстве случаев такие органы служат лишь для доставления преимуществ одному самцу над другим, потому что за достаточно долгий промежуток времени, менее одаренные самцы могли бы успешно спариваться с самками; а судя по строению самки, самцы эти во всех прочих отношениях одинаково хорошо приспособлены к обычному образу жизни. Так как в подобных случаях самцы прибрели свое нынешнее строение потому, что они лучше приспособлены к переживанию в борьбе за существование, но по той причине, что это преимущество было передано исключительно мужскому потомству[19], половой подбор должен был здесь играть роль. Важность этого различия и привела меня к обозначению этой формы подбора именем «половой». Далее, таким же образом, если главная польза, оказываемая самцу его хватательными органами[20], состоит в предупреждении побега самки до прибыли других самцов или до нападения с их стороны, то и эти органы должны были улучшиться путем полового подбора, т. е. благодаря преимуществу, приобретенному известными особями над своими соперниками. Но в большей части случаев этого рода, невозможно провести различия между действием естественного и полового подбора» (Дарвин, 1896: 197–198; курсив мой – Е.П.).
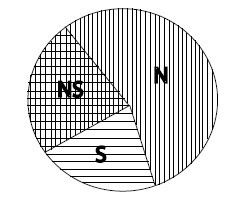
Рис. 1.1. Соотношение между естественным и половым отбором, как оно видится в доктрине полового отбора.
Из: Andersson, 1994.
Сказанное здесь, по-видимому, не кажется важным для Андерссона. Так или иначе, он ставит все точки над i в этих нечетких построениях Дарвина, облекая их в категорическую форму, которая приводится в виде графической схемы. Она показана на рис. 1.1, взятом из книги «Половой отбор». В подписи к нему сказано: «Соотношения между половым и естественным отбором. Внутри круга находятся все признаки (traits), находящиеся под действием (favored by) естественного отбора в природе. В левой половине круга содержатся те из них, которые связаны с конкуренцией из-за полового партнера (половой отбор NS и S). Некоторые признаки контролируются другим (other?) естественным отбором, как и половым отбором (NS) или только другим (?) естественным отбором (сектор N). Наконец, некоторые признаки подчиняются только половому отбору (сектор S). Этот сектор содержит набор признаков, имеющих отношение только к конкуренции между самцами. Это вторичные половые признаки, снижающие выживаемость (survival), которые Дарвин отнес к сфере действия «полового отбора». Заметьте, однако, что многие признаки контролируются как половым отбором, так и другими формами (?) естественного отбора. Относительная площадь разных секторов дана условно. Рассматриваются только те признаки, которые находятся под действием отбора в природных условиях, а не контролируются искусственным отбором или генетическим дрейфом» (Andersson, 1994: 8; курсив и знаки вопроса мои – Е.П.).
Легко видеть нечеткость и путанность предлагаемой здесь классификации форм отбора, что делает проведение границы между ними процедурой совершенно искусственной. Она не обоснована ничем, кроме чисто произвольного домысла. Это обстоятельство придает «теории» откровенный натурфилософский характер и тем самым ставит под удар все дальнейшие ее построения.
II. Межполовой и внутриполовой отбор. Внешнюю видимость того, что «половой отбор» есть некая реальная сущность, придает для непосвященных подразделение этой категории на два класса. Межполовой отбор осуществляется за счет селекции самками лучших самцов-производителей. Внутриполовой же идет, якобы, на почве конкуренции между самцами из-за самок. Он, как полагал еще Дарвин, должен быть особенно интенсивен у видов, у которых соотношение полов сильно сдвинуто в пользу самцов, что обостряет их конкуренцию из-за доступа к самкам. Второе понятие предложил Дж. Хаксли, который был склонен считать половой отбор лишь разновидностью естественного (Huxley, 1938а, б). К сожалению, от трактовок Хаксли, много более реалистичных, чем дарвиновские, в современной конструкции анализируемых взглядов сохранили одно лишь понятие внутриполового (интрасексуального) отбора (Huxley, 19386).
Сегодня теоретикам полового отбора кажется, что такое подразделение есть шаг вперед в сторону модернизации интуитивных построений Дарвина. Постулируя внутреннюю неоднородность полового отбора, они, как им кажется, уходят от чисто умозрительного (натурфилософского) взгляда на этот процесс, придавая ему видимость большей материальности и правдоподобия. В самом деле, если в явлении удается выявить хотя бы две разные составляющие, появляется возможность (в данном случае скорее иллюзорная) рассматривать его аналитически, не ограничиваясь одной лишь голой констатацией реальности этого явления, как это делал Дарвин.
III. Признаки самцов, способствующие их успеху у самок, снижают приспособленность их носителей. Идея, согласно которой самцы «обычно» окрашены ярче самок и несут вторичные половые признаки в виде всевозможных «украшений» и прочих эксцессивных структур (таких, например, как рога у многих копытных), стала главной в формулировании принципа полового отбора Дарвином. Проблема, с его точки зрения, состоит в том, что половой отбор, таким образом, противодействует естественному, который всегда должен работать в сторону повышения приспособления особей. В наши дни убеждение, согласно которому вторичные половые признаки самцов должны непременно снижать их выживаемость, нашла свое отражение в так называемом «принципе гандикапа» (Zahavi, 1975, 1993; Zahavi, Zahavi, 1997).
Суть этой идеи в том, что если уж самец смог выжить, несмотря на свою заметность, то его орнаментация означает для самки «честный» сигнал о его высоком качестве как производителя потомства.
Одно из отличий «честного» сигнала от всех прочих состоит, как полагают, в следующем. Все те структуры, которые рассматриваются в качестве украшений самца и при этом представлены в двойном наборе – справа и слева, как например, рога у копытных, должны, согласно теории, быть более или менее асимметричными. Это предположение базируется на том основании, что такие структуры есть результат интенсивного направленного[21] (strong directional) полового отбора. А тот должен, по мнению теоретиков этого направления, работать против генетических модификаторов, канализирующих развитие организма, в силу чего нарушается его стабильность. Именно это и должно приводить к неодинаковой скорости роста структур, обязанных действию полового отбора, в отличие от других, сугубо функционального характера (ноги, крылья и пр.[22]). Явление получило название «варьирующей асимметрия» (fluctuating asymmetry).
Варьирующей (флюктуирущей) она названа потому, что степень ее выраженности не одинакова у особей данной популяции (в том числе и самцов), причем отклонения от идеальной симметрии носят случайный характер (Parsons, 1990; подобнее см. раздел 10.3). Полагают, что у индивидов «высокого генетического качества» в период их онтогенеза некая буферная система должна работать против затрат (costs), тормозящих развитие ярко выраженных (elaborate) вторичных половых признаков, чего нет у генетически низкокачественных особей. В результате, высококачественные самцы будут иметь более крупные и более симметричные «честные» сигнальные структуры, а низкокачественные – меньшие по размерам и асимметричные (см., напр., Moller, Pomiankowski, 1993: 465). Все эти построения носят название «гипотезы честной сигнализации». В действительности, как нетрудно видеть, перед нами нанизывание друг на друга как минимум трех достаточно произвольных предположений, что вообще характерно для структуры рассматриваемой «теории».