Как было отмечено выше, каждый акт поведения есть актуализация систем разного уровня дифференциации, однако соотношение активных высоко- и низкодифференцированных систем, необходимых для успешной реализации поведенческого акта, различается для разных актов. Из единой концепции сознания и эмоций следует, что чем выше доля нейронов низкодифференцированных систем, активных в реализующемся поведении, тем выше интенсивность эмоций. В связи с этим можно предположить, что подавление активности нейронов, принадлежащих к высокодифференцированным системам, должно вести к относительному увеличению вклада систем низкой дифференциации и увеличению интенсивности эмоций как их характеристики.
Глава 2. Регрессия при острой алкогольной интоксикации
1. Мозговые механизмы регрессии
• Регрессия при введении алкоголя имеет в своей основе врéменное угнетение активности нейронов более новых дифференцированных систем – элементов субъективного опыта, т. е. врéменную дедифференциацию.
• Дедифференциация – обратимое относительное увеличение представленности в актуализированном опыте менее дифференцированных систем.
• Увеличение относительного вклада рано сформированных менее дифференцированных систем в обеспечение поведения связано с увеличением интенсивности эмоций.
• Алкоголь сильнее влияет на новый опыт, чем на старый, и на более сложное поведение, чем на простое.
В многочисленных экспериментах, проведенных нами, было показано, что под влиянием алкоголя происходит относительное увеличение вклада реализации систем низкой дифференциации в обеспечение поведения как у человека, так и у животных. Его острое введение (этанол; 1 г/кг) животным вызывает достоверное и обратимое уменьшение числа активных в поведении нейронов мозга, принадлежащих к наиболее новым и дифференцированным системам (Alexandrov et al., 1990а, 1991, 2013; позднее сходные данные были получены и другими авторами; см. ссылки в Alexandrov et al., 2013). Например, в лимбической коре их относительное число уменьшается в 2,5 раза.
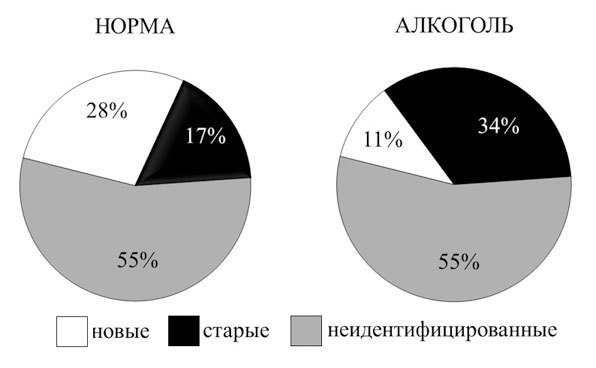
Рис. 1. Процент нейронов разной специализации в лимбической коре кролика в норме (инъекция физиологического раствора) и после введения алкоголя
Относительное число нейронов с неидентифицированной специализацией (н/и; не удается выявить постоянную связь изменения частоты их активности с актами анализируемого поведения) не меняется. В то же время видно уменьшение (с 28 % до 11 %) относительного числа активных нейронов, принадлежащих к наиболее новым (Нов) дифференцированным системам, сформированным на последнем, непосредственно предшествующем регистрации активности нейронов этапе индивидуального развития (обучение животных инструментальному пищедобывательному поведению), и увеличение (с 17 % до 34 %) относительного числа активных нейронов, принадлежащих к системам, сформированным на ранних этапах индивидуального развития («старые»; Ст). При этом достоверно возрастало число ошибок в поведении. Специальные подсчеты (произведенные при учете числа активных нейронов, обнаруженных при движении микроэлектрода сквозь толщу коры) показывают, что отмеченное выше изменение относительного числа нейронов обусловлено достоверным уменьшением абсолютного числа активных нейронов новых систем после введения алкоголя. После завершения действия алкоголя число активных нейронов новых систем возвращается к норме.
Более выраженное угнетающее влияние (как острое, так и хроническое – Alexandrov et al., 2001, 2013) алкоголя на новые сравнительно более дифференцированные системы (согласующееся с принципом Рибо – Джексона – Ribot, 1901) было выявлено нами также при сопоставлении эффектов этанола на медленные электроэнцефалографические потенциалы мозга, регистрируемые у птенцов при актуализации систем, обеспечивающих поведенческие акты, которые последовательно формируются на самых ранних стадиях онтогенеза. Было выявлено, что при введении алкоголя реализация вновь сформированного более сложного (оборонительного) поведения сопровождается более выраженным избирательным угнетением мозговой активности (уменьшением амплитуды ЭЭГ-потенциалов, вызванных значимым сигналом) по сравнению с ранее сформированным (пищедобывательным) поведением (Alexandrov, Alexandrov, 1993).
Мы исследовали влияние острого введения алкоголя на мозговую активность незрелорождающихся птенцов мухоловки-пеструшки через 4–8 дней после их вылупления. Птенцам предъявляли звуки (посылки чистых тонов), связанные с реализацией естественного поведения (2,0 и 5,0 кГц; пищедобывательное и оборонительное поведение, соответственно), а также контрольный звук (тон), не имеющий поведенческой значимости для птенцов данного возраста (3,0 кГц). Регистрировали вызванные ЭЭГ-потенциалы при предъявлении этих звуков на последовательных стадиях формирования и последующего увеличения дифференцированности поведения. После регистрации вызванных потенциалов птенцу давали личинку мучного хрущака, в которую непосредственно перед этим вводили этиловый спирт (96 %) из расчета дозы 1–1,2 г/кг массы тела. Острое введение алкоголя птенцам мухоловки-пеструшки приводило к уменьшению амплитуды слуховых вызванных потенциалов, возникающих в поле L каудального неостриатума в ответ на предъявление звуков, связанных с пищевым и оборонительным поведением, но не контрольного звука (рисунок 2). Анализ динамики амплитудно-временных показателей (коэффициента зрелости) вызванных потенциалов при введении алкоголя позволил сделать вывод о том, что имеет место сдвиг показателей в сторону меньшей зрелости.
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.