В этой связи время развития и протяженность репродуктивной фазы столь же важны, как и длительность периода старения. По крайней мере у млекопитающих существует, по-видимому, положительная корреляция между временем, необходимым для достижения половой зрелости, и максимальной продолжительностью жизни [6, 57] (табл. 1.1). У вида Homo sapiens, который среди млекопитающих живет дольше всех, самый длинный период развития; у индийского слона, лошади и крупного рогатого скота период развития соответственно короче. Однако существуют исключения, особенно среди мелких животных. Эти исключения могут быть примерами адаптивных изменений, которые возникают в особых условиях обитания. У всех млекопитающих, а также у большинства других видов животных репродуктивный период занимает значительную часть жизни и сменяется пострепродуктивным периодом. Однако среди беспозвоночных и низших животных имеются случаи смерти вскоре после единственного акта размножения; таковы тихоокеанский лосось (Onchorhynchus), угорь, речная минога, небольшие однолетние рыбы и осьминог (Octopus). Вероятно, у этих видов воспроизводство вызывает резкое снижение некоторых важных факторов, которые не восстанавливаются, хотя и являются жизненно необходимыми.
В связи с этим встает вопрос о скорости репродукции, или плодовитости, о продолжительности репродуктивного периода и о продолжительности жизни после достижения половой зрелости. Мелкие млекопитающие типа грызунов, которые обладают высокой плодовитостью, имеют короткую репродуктивную фазу, а также укороченный пострепродуктивный период по сравнению с такими крупными млекопитающими, как слон, лошадь, домашний рогатый скот и человек, у которых скорость воспроизводства много ниже. Вероятно, чем быстрее воспроизводство, тем быстрее потеря определенных важных факторов и тем быстрее развитие разрушительных изменений. Однако черепахи откладывают много яиц, но тем не менее долго живут [116]. Матки общественных насекомых, пчел и термитов, которые отличаются большой плодовитостью, также имеют большую продолжительность жизни.
С вышесказанным связано наблюдение, что среди млекопитающих постепенное увеличение времени достижения половой зрелости происходит параллельно с увеличением максимальной продолжительности жизни. Это особенно очевидно для человекообразных. Во время их эволюции за последние 3 млн. лет максимальная продолжительность их жизни выросла почти в 2 раза со скоростью прироста около 14 лет на 100000 лет [23]. Это возможно только при изменениях в геноме типа мутаций и перегруппировки генов. Вызывают ли генетические изменения, результатом которых является увеличение продолжительности жизни, сокращение плодовитости, наблюдаемое у этих долгоживущих млекопитающих, или удлинение периода размножения приводит к увеличению продолжительности жизни, неизвестно.
Теперь коснемся проблемы начала и продолжительности старения. Если начало репродуктивного периода определяется для каждого вида более или менее отчетливо, то этого нельзя сказать о начале старения. Если принять время прекращения репродукции за начало старения (хотя установлено, что функциональная способность практически всех органов начинает снижаться гораздо раньше и что скорость репродукции замедляется задолго до ее прекращения), то следует отметить, что у высших млекопитающих период старения увеличивался параллельно с увеличением репродуктивного периода. Фаза старения не дает особых преимуществ, когда речь идет о выживании, сохранении и эволюции вида, и тем не менее эта фаза увеличилась. Не происходит ли это потому, что низкая скорость репродукции оказывает меньшее вредное воздействие на другие жизненные функции, которые, таким образом, дольше сохраняются? К сожалению, достоверных данных, касающихся пострепродуктивной фазы у животных, недостаточно для установления связи между периодами развития и репродукции и периодом старения.
Если даже рассматривать геном в качестве того первичного участка, где возможны перемены после достижения животным половой зрелости, необходимо иметь в виду, что изменения внеклеточного окружения, связанные с температурой, питанием и т. д., существенно влияют на функцию генома. Поэтому у представителей различных видов и популяций наблюдаются колебания во времени начала старения, его скорости и продолжительности пострепродуктивного периода.
Полученные к настоящему времени данные указывают на то, что старение имеет генетическую основу. Рядом авторов разработаны теории, в которых делаются попытки объяснить старение на уровне генома. Ниже мы обсудим теории только этого типа.
Теория соматических мутации
Росс и Скотт [104] первыми сообщили, что крысы, подвергнутые тотальному облучению, слишком слабому, чтобы вызвать какие-либо острые изменения, погибают раньше, чем необлученные контрольные животные. Затем последовали сообщения о том, что у облученных грызунов [26, 44, 108] и людей [128] симптомы старения, и смертность были такими же, как у интактных особей; было отмечено, однако, что частота опухолей у первых была выше. Поэтому предположили, что облучение вызывает ускорение процесса старения. Основываясь на этих данных, Сцилард [122, 123] предложил для объяснения старения "теорию соматических мутаций", согласна которой мутации, возникающие беспорядочно и самопроизвольно, разрушают гены и хромосомы постмитотических клеток в течение жизни организма, постоянно повышая мутационный груз. При увеличении числа мутаций и потере функциональных генов наблюдается снижение синтеза функциональных белков. Смерть клетки наступает тогда, когда мутационный груз превышает критический уровень. В результате число постмитотических клеток уменьшается, а общая функциональная активность организма снижается.
Эта теория была проверена в опытах с мышами Стивенсоном совместно с Кёртисом [118] и Кёртисом [20, 21]. Мышей облучали дозой 400 рад или им вводили химические мутагены типа азотистых аналогов иприта (0,125 мг) и затем определяли их выживание. В регенерирующей печени молодых и старых мышей исследовали также частоту хромосомных аберраций типа нерасхождения сестринских хроматид в митозе и хромосомных разрывов. У мышей из линий с различной продолжительностью жизни вызывали частичный некроз печени введением четыреххлористого углерода и подсчитывали число хромосомных аберраций после фиксации в метафазе под действием колхицина.
Было показано, что а) сокращение жизни после рентгеновского облучения зависит от дозы, б) в регенерирующих клетках печени облученных мышей повышена частота хромосомных аберраций (рис. 9.1), в) азотистые аналоги иприта не оказывают влияния ни на продолжительность жизни, ни на частоту хромосомных аберраций, г) в клетках печени короткоживущих мышей линии A/HEJ (395 дней) накапливается больше хромосомных аберраций, чем у долгоживущих мышей линии C57BL/6J (600 дней; рис. 9.2) [19]. Кёртис [20] далее предположил, что проникающая радиация повреждает постмитотические клетки больше, чем премитотические. В первых мутационные эффекты аккумулируются и клетка не может устранить их, так как она не делится. Во втором случае поврежденная клетка элиминируется и замещается неповрежденной клеткой. Клетки зародышевого пути более устойчивы к повреждению хромосом, благодаря чему возможно сохранение вида.
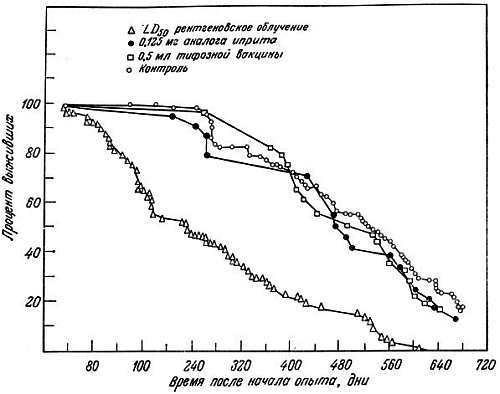
Рис. 9.1. Кривые выживания мышей после различных воздействий, начиная с 2-месячного возраста [20]. Кривые начинаются через 30 дней после воздействия — срока, достаточного для того, чтобы не учитывать внезапную гибель. Они показывают, что однократные массивные, но не смертельные дозы ядовитых химических веществ не уменьшают продолжительности жизни, тогда как единичные массивные, но не смертельные дозы рентгеновского облучения дают заметный эффект