Старение клеток после трансплантации
У насекомых с полным превращением, например у дрозофилы, на определенных участках эмбриона расположены группы специализированных клеток — имагинальные диски. Клетки каждого диска предназначены для дифференцировки специфического типа, т. е. превращения в определенный орган. Когда имагинальный диск эмбриона пересаживают в полость тела личинки того же вида, эти клетки пролиферируют, но остаются в недифференцированном состоянии до образования куколки. В период окукливания они дифференцируются и превращаются в определенную структуру. После дифференцировки эти клетки не пролиферируют. Если же имагинальный диск пересадить в тело взрослой особи того же вида, то клетки диска делятся, но не дифференцируются до конца жизни хозяина. Если диск извлечь в любом периоде жизни хозяина, разделить на фрагменты и пересадить взрослым особям, то клетки продолжают делиться у реципиента, не дифференцируясь. Используя метод серийных трансплантаций, Хейдорн [20] показал, что клетки имагинального диска сохраняют пролиферативную активность длительное время, во много раз превышающее максимальную продолжительность жизни насекомого. Эти исследования доказали, что, по крайней мере у насекомых, клетки пролиферируют в определенных условиях в течение времени, которое существенно больше продолжительности жизни хозяина. Однако неизвестно, замедляется ли постепенно пролиферативная активность или нет. Имагинальные диски имеются только у насекомых; так как у позвоночных сравнимые эмбриональные структуры отсутствуют, значение этих исследований для старения высших животных остается проблематичным.
С целью изучения процессов старения клеток и тканей производили пересадку тканей мышей одного возраста реципиенту (хозяину) другого возраста. Эти исследования выполнялись на инбредных и сингенных мышах, чтобы по возможности избежать несовместимости тканей. Крои [31, 32] серийно пересаживал кожу тела, уха и хвоста мышей-родителей молодым гибридам F1. Когда хозяин F1 старел, трансплантаты отделяли и пересаживали мышам F2. Такие последовательные переносы лоскутов кожи показали, что ее жизнеспособность и пролиферативный потенциал клеток сохраняются в течение 7–8 лет, что значительно превышает продолжительность жизни мыши. Однако после 5–6 пассажей трансплантат уменьшался в размере, с каждым последующим переносом пролиферация клеток снижалась и в конце концов ткань дегенерировала. Можно сделать вывод, что клетки сохраняют свою жизнеспособность в течение времени, превышающего продолжительность жизни хозяина, а ограничение пролиферативной активности зависит от условий внеклеточного окружения.
Влияние возраста донора на пересаженную ткань было изучено путем трансплантации кожи старой и молодой (возраст 680 и 74 дня) мыши 74-дневному животному [32]. Старая кожа отмирала у молодой мыши через 199–200 дней, но молодая ткань сохранялась дольше, до 300 дней. Хортон [29] пересаживал кожу старых (30–32 мес) мышей молодым (3–4 мес) реципиентам. Трансплантаты переживали более 4 лет, что значительно больше максимальной продолжительности жизни мышей. Следует отметить, что скорость деления клеток трансплантированной ткани была снижена. Это может быть одной из причин ее более длительного выживания.
Клетки кроветворной системы сохраняют высокую пролиферативную способность in vivo, и, по-видимому, это свойство не подвержено возрастным изменениям. Кроветворная ткань содержит стволовые клетки, или клетки-предшественники. Полагают, что каждая стволовая клетка делится, образуя одну стволовую клетку — потомка и дифференцированную клетку. Последняя начинает экспоненциально делиться с образованием клеток, которые имеют малую продолжительность жизни. Стволовые клетки имеют большую продолжительность жизни и сохраняют способность к пролиферации в течение всей жизни животного. Способность этих клеток пролиферировать была исследована посредством серийных пересадок молодым реципиентам, чья кроветворная система была разрушена массивным облучением всего тела (~800 рад). Когда реципиенты старели, трансплантированные кроветворные клетки переносили более молодым облученным реципиентам. Было показано, что защита от последствий облучения, создаваемая пересаженными клетками у реципиента, с каждой пересадкой ослаблялась [1,6]. Это может свидетельствовать об ограниченном пролиферативном потенциале стволовых клеток. В другом исследовании облученным мышам внутривенно вводили живые кроветворные клетки. Через 9-14 дней стволовые клетки образовывали колонии в селезенке хозяина. Каждая колония: развивалась из одной стволовой клетки. Клетки этих колоний были затем серийно пересажены более молодым облученным реципиентам (рис. 8.4). Результаты показали, что число колоний, образующихся в селезенке, снижается с каждой трансплантацией [33, 34]. Даже если согласиться с тем, что пролиферативная активность клеток кроветворной системы снижается после определенного возраста, оставалось необъяснимым влияние больших доз радиации на старение кроветворной системы и организма.
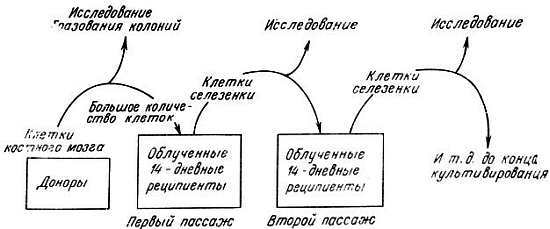
Рис. 8.4. Исследование колониеобразующей активности клеток костного мозга методом серийных трансплантаций облученным реципиентам [44]
Ситуация стала более ясной, когда в качестве реципиентов, кроветворных клеток вместо облученных мышей использовали, животных с генетически детерминированной анемией [21, 22]. Клетки молодых и старых доноров излечивают анемию и одинаково хорошо размножаются в течение 36 мес при четырех последовательных трансплантациях. Однако наблюдается постепенное снижение колониеобразующей активности стволовых клеток, которая регистрируется по числу колоний в селезенке реципиента, причем снижение больше выражено для клеток старых доноров. По-видимому, можно сделать вывод, что кроветворные клетки способны лишь к ограниченному числу делений, хотя они могут делиться в течение времени, превышающего продолжительность жизни мыши.
В другой работе Уильямсон и Асконас [52] иммунизировали мышей-доноров иммуноглобулином G (быка), конъюгированным с динитрофенолом. Иммуноглобулин, вырабатываемый у мышей к этому антигену, очень специфичен, и его можно легко обнаружить. Клетки костного мозга иммунизированных мышей затем трансплантировали реципиентам, иммунная система которых была разрушена интенсивным облучением (рис. 8.4). У реципиентов исследовали продукцию данного иммуноглобулина. Производили серийные пересадки клеток костного мозга от одного поколения мышей другому. После четырех трансплантаций выработка иммуноглобулинов снижалась вследствие постепенного уменьшения скорости деления с каждой последующей пересадкой, хотя кроветворные клетки были функционально активными в течение периода времени, превышающего максимальную продолжительность жизни мышей. В экспериментах Харрисона [21, 22] и Уильям сон а и Асконаса [52] не было установлено, какое влияние на пролиферацию оказывает сама процедура трансплантации и какие факторы в норме способствуют снижению потенциала деления кроветворных клеток.
Опыты по изучению влияния возраста донора на пролиферативную способность кроветворных клеток дали противоречивые результаты. Харрисон и Даблдей [23] иммунизировали молодых и старых мышей эритроцитами барана (SRBC). Клетки костного мозга и селезенки затем серийно пересаживали молодым летально облученным реципиентам и оценивали колониеобразующую активность стволовых клеток в селезенке. Оказалось, что клетки мышей обоих возрастав имеют почти одинаковую способность к образованию колоний. Это противоречит данным Макинодана и др. [36] и Уильямсона и Асконаса [52], которые сообщили об ослаблении иммунной компетентности у мышей с возрастом. Возможно, что эти различия объясняются неодинаковой длительностью интервалов между пересадками.