Были предприняты попытки установить взаимосвязь между продолжительностью жизни вида и потенциалом деления фибробластов, и хотя были изучены лишь немногие виды с разной продолжительностью жизни, результаты указывают на наличие положительной корреляции между этими величинами (табл. 8.2). Фибробласты эмбриона галапагосской черепахи, наиболее долгоживущего вида среди изученных, обладают и самым высоким потенциалом деления, тогда как для мыши, чья продолжительность жизни не превышает трех лет, эта величина значительно меньше.
Таблица 8.2. Максимальная продолжительность жизни культивируемых нормальных фибробластов эмбриона человека и животных [26]

Для ответа на вопрос, вызваны ли старение и смерть клеток накоплением какого-либо цитоплазматического фактора, был использован метод гибридизации соматических клеток [53]. Гибридизация старых и молодых клеток была нежелательна, так как она привела бы к образованию гибридов с двойным набором хромосом. Удалением ядра из легочных фибробластов человека, находившихся во II и III фазах роста, получали цитопласты. Затем происходило их слияние с интактными фибробластами раннего и позднего пассажей следующим образом: молодые цитопласты — с молодыми клетками, старые цитопласты — с молодыми клетками, молодые цитопласты — со старыми клетками и старые цитопласты — со старыми клетками. Число удвоений популяции первых двух гибридных линий составило около 20, а двух последних — около 5. Отсюда следует, что цитоплазма, вероятно, не содержит никакого фактора, который влияет на потенциал деления, т. е. пролиферативная активность клеток определяется на уровне ядра или генома. Полученные результаты необходимо подтвердить в дальнейших исследованиях.
Таким образом, клетки стареют как in vivo, так и in vitro. Существует, однако, большая вариабельность продолжительности их жизни и числа делений. Клетки типа нейронов или скелетной и сердечной мышц прекращают деление in vivo на ранних стадиях развития и становятся постмитотическими. Было бы интересно узнать, сколько раз они подвергаются делению, прежде чем достигают постмитотической стадии. Спустя некоторое время эти постмитотические клетки стареют и погибают. Таким образом, эти клетки сначала прекращают деление, а затем подвергаются старению. Клетки костного мозга, эпителия и печени сохраняют пролиферативную активность в течение всей жизни, хотя она постепенно падает в старческом возрасте. Когда эти клетки культивируют iv vitro, они прекращают делиться и через некоторое время стареют. Очевидно, эти клетки обладают способностью к размножению, которая во много раз превышает аналогичную способность нейронов и мышечных клеток. Важно изучить клетки эмбриона тотчас после того, как они прошли дифференцировку и превратились в клетки специфических типов, чтобы увидеть, имеют ли степень и тип дифференцировки какую-либо связь с пролиферативной активностью и природой постмитотического состояния. Такие исследования клеток различных типов, начиная с самых ранних стадий дифференцировки и до прекращения деления, могут дать ключевую информацию о причинах прекращения деления.
Возникает вопрос, действительно ли те клетки, которые сохраняют способность делиться в течение всей жизни, не стареют? Действительно ли они обновляются всякий раз, когда делятся? Можно ли считать две дочерние клетки полностью идентичными не только в отношении содержания ДНК, но и родительских органелл? Известно, что во время созревания сперматиды наблюдается неадекватное деление.
Вернет [3] предположил, что клетки одного типа могут быть более ответственны за старение, чем другие. По его мнению клетки тимуса и тимус-зависимые клетки, которые подвержены ослаблению пролиферации по типу фазы III, определяют темп старения. Вернет — сторонник той точки зрения, что изменения в этих клетках ответственны за аутоиммунные заболевания, которые чаще встречаются в пожилом возрасте. Эта концепция основана на представлении, что причиной старения являются соматические мутации, однако доказательств для нее пока нет.
Если старение — внутреннее свойство клеток, тогда оно должно быть присуще и одноклеточным организмам. Амебы проходят ограниченное число делений и живут ограниченное время, если их содержать на неполноценном рационе. Они продолжают расти и размножаться непрерывно только в том случае, если питание обильно, чего обычно не бывает в естественных условиях [38]. Многие клоны парамеций [46, 47] и аскомицетов [41] имеют ограниченную пролиферативную активность.
Старение клеток in vivo и после трансплантации
Клетки некоторых типов стареют in vivo после немногочисленных делений, другие — после большого числа делений. Различия зависят от степени дифференцировки клеток, их окружения и питания. Ограниченная жизнеспособность клеток может быть следствием снижения функциональной активности и старения организма в целом. Нельзя ли избежать старения и надолго сохранить клетки живыми путем переноса их молодому хозяину? Вместе с тем, если клетки организма постоянно замещать клетками более молодого индивидуума, не сделает ли это его жизнь более продолжительной или бесконечной? Трансплантация клеток молодому хозяину обеспечивает им естественное физиологическое окружение, которое нельзя создать при культивировании вне организма. Поэтому опыты по трансплантации позволяют отчасти отклонить упреки по поводу неестественного окружения клетки при культивирований in vitro и неприменимости полученных этим способом результатов к процессу старения организма.
Старение клеток in vivo
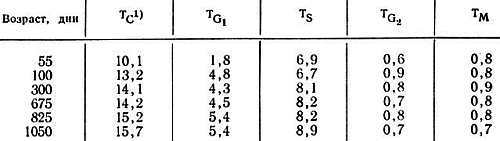
Ранние работы по старению клеток in vivo были проведены на интактных объектах. Уайтли и Хортом [50] нашли, что число митотических фигур на единицу поверхности клеток эпидермиса уха мыши составило 30, 15 и 2–6 в возрасте 9 дней, 3 дней и 33–36 мес (старые мыши) соответственно. Трашер и Гройлих [49], изучая радиоавтографически пролиферативную активность клеток двенадцатиперстной кишки мышей в возрасте 10, 30–70, 380–390 и 579–638 дней (старые мыши), обнаружили, что продолжительность клеточного цикла с возрастом увеличивается (табл. 8.3). Преимущественно увеличивалась продолжительность G1-фазы, тогда как фаза синтеза ДНК (S-фаза) не изменялась. Эти исследования ясно показывают, что, хотя клетки некоторых типов сохраняют способность к пролиферации вплоть до очень старого возраста, их активность существенно снижается.
Таблица 8.3. Время митотического цикла клеток обкладочного эпителия крипт тонкого кишечника мышей линии BCF1 в различном возрасте [2]
1) TC, TG1, TS, TG2 и ТM — соответственно продолжительность (в часах) клеточного цикла, фаз G1, S, G2 и митоза.
Такое явление, как смерть клетки, встречается не только в старческом возрасте или в фазе III при культивировании, клетки гибнут и в период раннего развития. Жабры и хвост головастика, предпочка и первичная почка высших позвоночных элиминируются в период раннего развития вследствие отмирания клеток этих органов [42]. Образование и формирование пальцев конечностей у позвоночных также сопровождается процессами гибели и резорбции клеток [51]. При метаморфозе, т. е. превращении личинки насекомого во взрослую особь, наблюдается массовое отмирание клеток органов личинки. Следовательно, гибель клеток в период развития необходима для образования совершенных и функционально активных органов взрослых особей, она своевременна и наступает на соответствующих стадиях развития, т. е. и здесь однажды сформированные клетки стареют и умирают.