Для абсолютного большинства характеристик животного, определяемых как "признаки", почти невозможно вычленить отдельные локусы или, во всяком случае, определить действие отдельных генов по внешним показателям. Рассмотрим один из количественных признаков - например, длину хвоста.
Предположим, что итоговая длина определяется генами неких локусов X, Y, Z. При этом гомозиготы XX YY ZZ будут иметь наиболее короткие хвосты, гомозиготы xx yy zz - самые длинные. Те кошки, у которых одна пара генов будет находиться в доминантном гомозиготном состоянии, а другие - в рецессивном, будут показывать промежуточную длину хвоста
(Xx yy zz, xx YY zz и т.д.). Что же касается гетерозигот, то проявление у них признака будет зависеть от степени доминирования аллелей каждой пары. При полном доминировании гетерозиготы Xx Yy Zz будут неотличимы от XX YY ZZ, при неполном - опять-таки покажут промежуточные характеристики.
Мало того, гены полигенного комплекса могут отличаться разной. активностью. Например, гены Х-локуса оказывают большее влияние на длину хвоста, чем гены локуса Z. Тогда особи хх уу ZZ окажутся более длиннохвостыми,
чем XX yyzz. Так в крайне упрощенном виде можно представить тип аддитивной, или кумулятивной, полигении. Есть и другие ее типы, например, комплементарная. Признак, определяемый таким образом, будет выражен внешне только в случае, когда каждый из генов полигенного комплекса находится в определенном аллельном состоянии. То есть у кошки генотипа UU Yy zz некий признак выражен, а у имеющих прочие варианты - uu YY zz, UU yy Zz, UU Yy ZZ - нет.
Строго говоря, таких признаков, за которые отвечал бы один и только один ген, практически нет. Даже на те признаки, которые, казалось бы, имеют явно моногенную природу, оказывает влияние; генетический фон — гены других локусов.
Отчасти вариации в проявлении наследственных признаков объясняются влиянием окружающей среды. Наследуется ведь не просто признак в полном его выражении, а, скорее, возможность выражения признака в некоторых пределах, или, иначе говоря, норма реакции. Однако выраженность признака определяется не только условиями жизни организма. Например, степень ослабления окраса, вызванная рецессивным аллелем гена Dilution dd, будет косвенно зависеть от генов, отвечающих за текстуру шерсти. Такие гены, влияющие на проявления аллелей других генов, называют модификаторами.
Для того чтобы охарактеризовать их влияние на проявление того или иного признака, пользуются понятиями экспрессивности и пенетрантности. Экспрессивность - это степень проявления признака. Если она сильно зависит от действия модификаторов, говорят о вариабильной экспрессивности гена, если же нет — о высокой или, стабильной. Пенетрантность определяют как часть особей, внешне проявляющих признак, от числа всех особей, несущих аллели, определяющие этот признак. Пенетрантность может быть полной -например, все кошки, несущие аллель L, имеют короткую шерсть; или неполной - из числа кошек, несущих ген полидактилии (избыточного количества пальцев) Pd, проявляют эту аномалию меньше половины. При значительной выборке пенетрантность может быть вычислена в процентах. Для известного гена доминантного белого окраса White пенетрантность в отношении окраса почти 100%, голубоглазости - около 70% и глухоты - около 40%. Кстати, и экспрессивность, и пенетрантность не являются совсем уж неизменными характеристиками генов. Они значительно варьируют в каждой конкретной кошачьей популяции.
РЕГУЛЯЦИЯ РАБОТЫ ГЕНА
Чем же обусловлена такая сложная картина наследования, столь отличающаяся от простых законов Менделя? Прежде всего - гигантской системой регуляции работы гена. Далеко не вся ДНК организма состоит исключительно из генов. Значительная ее часть "молчит", то есть не считывается на матричную РНК.
Даже внутри участка, относящегося к какому-либо гену, не вся информация представляет собой код для синтеза белка. Однако она содержит ряд регуляторов деятельности гена, которые обеспечивают возможность считывания информации, ускоряют или замедляют этот процесс.
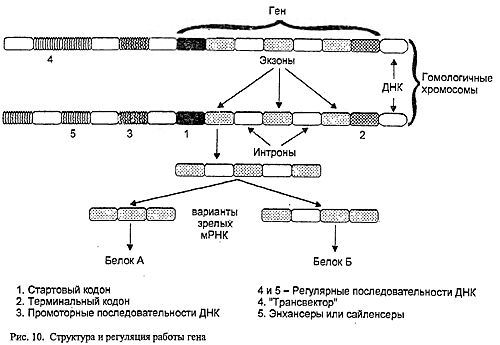
Перед каждым геном находится особая нуклеотидная последовательность - промотор, которая служит сигналом к началу переписывания информации на РНК (рис.10).
Изменения в этой последовательности будут вести или к снижению интенсивности процесса считывания информации на РНК (транскрипции), или к его прекращению. Управлять активностью транскрипции внутри клетки могут и так называемые операторы - другие нуклеотидные последовательности, которые взаимодействуют с белками, блокирующими "считывание".
Благодаря этому процессу происходит "выключение" гена, когда кодируемого им белка в клетке уже достаточно для выполнения ее функций. (Регуляция работы гена лучше изучена у вирусов и бактерий, отличающихся простой организацией генома. У млекопитающих сохраняются эти типы регуляции, но обеспечивающие их структуры могут быть более сложными.)
Но нельзя забывать о том, что у высших организмов, помимо рёгуляторных процессов, управляющих жизненным циклом одной клетки, протекают и Те реакции, которые определяют жизнедеятельность и развитие организма в целом. Не в каждой клетке в одно и то же время обязательно работают все гены.
Их "включение" и "выключение" при развитии организма обусловлено действием множества ферментных и неферментных белков, гормонов и других биологически активных веществ.
Вообще спектр даже только одних белков, влияющих на генную активность, весьма разнообразен. Как было отмечено, "рабочим" является только одно, "раскрученное", состояние ДНК (конформация), для обеспечения которого необходим ряд структурных белков и ферменты-топоизомеразы. Все эти регуляторные белки, естественно, не появляются ниоткуда, а сами, в свою очередь, являются продуктами соответствующих генов.
Концепция классической генетики "один ген - один признак" трансформировалась впоследствии в "один ген - один белок". Однако и его положение оказалось неверным. Внутри многих генов были обнаружены участки двух типов - первые считывались на РНК и служили основой для синтеза белка (так называемые экзоны), вторые, хотя и считывались, но в процессе созревания этой РНК выпадали (интроны). Оставшиеся фрагменты-экзоны объединялись в итоговую цепь РНК. Казалось бы, интроны - структуры совершенно бессмысленные. Но "впоследствии было обнаружено, что отрезки РНК, образовавшиеся при с вырезании "вставок", могут служить матрицей для синтеза белка и по отдельности, и соединяться в цепочку различными способами. А это введет уже к образованию различных белков. То есть из исходного гена может образоваться несколько разных белков. Процессы созревания матричной РНК, разумеется, имеют свои регуляторные механизмы...
Кроме того, на активность генов могут влиять не только расположенные рядом с ним последовательности ДНК, но и находящиеся на значительном удалении от него на той же хромосоме или даже на гомологичной. Последовательности ДНК, увеличивающие синтез РНК (а значит, и белка) в сотни и тысячи раз, носят название энхансеров, а уменьшающие - сайленсеров. Процесс регуляции активности генов, расположенных на одной хромосоме, с помощью последовательностей ДНК ее гомолога, носит название трансвекции. Если методом генной инженерии вырезать последовательность-энхансер и вставить ее в нить ДНК в соответствующем положении к интересующему вас рецессивному аллелю, его активность и количество образуемого им белка превзойдет доминантный гомолог, результатом чего может быть промежуточное проявление признаков независимо от их доминантности-рецессивности и даже обратное соотношение, когда организм будет проявлять признак, противоположный ожидаемому (доминантному).