Известно, что частота мутаций иногда превышает естественный уровень на несколько порядков. Прежде всего, такое повышение может быть обусловлено мутагенами — химическими веществами, лекарственными препаратами, радиацией, ультрафиолетовым излучением. Но все-таки далеко не все мутанты — "дети Чернобыля". Зачастую повышение мутабильности генов связано не с загрязнением окружающей среды, а с совершенно иными факторами. Даже самый процесс одомашнивания животных может являться мутагенным фактором.
Это было показано в знаменитых опытах академика Беляева по одомашниванию лисиц. В каждом поколении лис исследователи отбирали для разведения животных, не проявлявших агрессивности по отношению к людям. Через несколько поколений этого отбора в популяции домашних лисиц появились такие признаки, как белая пегость, вислоухость, закрученный в колечко хвост.
Всякий организм представляет собой целостную систему, все физиологические процессы которой находятся в динамическом равновесии. Отбор животных по поведению одновременно явился отбором по определенному гормональному спектру и биохимии нервной системы. Генетикам известно, что направленный, движущий отбор по, каким-то признакам (не только по поведению) способен спровоцировать появление в популяции неспецифических (то есть не связанных с селективным признаком) мутаций. Правда, показана эта закономерность на плодовой мушке, а не на кошке, но не исключено, что окажется справедлива и для последней. Эти неожиданные эффекты возникают потому, что всякий признак контролируется не одним геном, а целостной, находящейся в равновесии генной системой. И когда в результате движущего отбора возрастает выраженность признака, это означает, что изменяются дозы и активности всех генов системы. А такое изменение грозит дисбалансом в работе всего генома, нарушаются взаимосвязи генных комплексов и возникают "ошибки" генетического кода.
Теоретически вспышка мутабильности такого рода возможна в каком-либо крупном питомнике, работающем с использованием инбридинга и при интенсивном отборе.
Известны, впрочем, и другие случаи спонтанного увеличения частоты мутаций — так называемые "мутационные взрывы". Иногда этим выражением характеризуют ситуацию, сложившуюся в фелинологическом мире в 50-60е годы.
Действительно, эти годы отмечены появлением нескольких мутаций, давших начало новым породам: корниш- и девон-рексы, балинезы и сомали, канадские сфинксы и шотландские фолды.
Понятие мутационного взрыва разработано опять-таки в применении к излюбленному объекту генетиков, плодовой мушке дрозофиле, и неизвестно еще, можно ли его отнести к млекопитающим. "Взрывы" эти возникают, как правило, в строго определенных генах и связаны с деятельностью мобильных, то есть способных к перемещению между хромосомами, фрагментов ДНК. Такие элементы заключают в себе информацию, позволяющую им самопроизвольно "вырезаться" из одного участка хромосомы и "вставлять" себя в другой. Встраиваясь в области, прилегающие к какому-нибудь гену, они провоцируют изменения в нем. Причины мутационных взрывов (возникающих, кстати, почти одновременно в нескольких удаленных друг от друга популяциях) до сих пор точно не изучены. Многие исследователи полагают, что такие вспышки мутабильности возникают вследствие вирусных заболеваний (вирусы, как и мобильные элементы ДНК, способны встраиваться в геном организма). Трудно сказать, была ли ситуация с породообразованием 50-х гг. "мутационным взрывом" или только взрывом интереса к кошачьим мутациям. Нам неизвестно, была ли в 50-е годы отмечена массовая вспышка вирусного заболевания, охватившая по крайней мере два континента. Да и какой-либо закономерности в том, какие именно гены мутировали у кошек в те годы, вроде бы не наблюдалось.
Как уже было сказано, существует ряд принципиально важных для организма генов, обеспечивающих основные процессы жизнедеятельности - такие гены наиболее консервативны, то есть представлены наибольшим числом копий. Это многократное дублирование позволяет части генных копий накапливать мутационные изменения и передавать их по наследству. Наличие вредных мутаций в отдельных копиях генов никак не сказывается на здоровье всего организма. Если же в них возникнет новая, более благоприятная, чем прежняя, комбинация, эта изменения копия гена с течением поколений станет основной в геноме вида. Что же касается не столь значимых для жизнедеятельности организма генов, то мутационный процесс постоянно создает их новые вариации, нередко закрепляющиеся, тем более при искусственном отборе.
Вариации одного и того же гена носят название аллелей. Образуемые разными аллелями одного гена белки частично отличаются по своей структуре и функции, но определяют в конечном итоге проявление (или отсутствие) одного и того же признака. Итак, реально гены существуют как аллельные варианты.
Поскольку за счет парности хромосом каждый ген представлен в организме, так сказать, дважды, его аллельные варианты, занимающие идентичные локусы в этих хромосомах, могут или совпадать, или быть различными. В первом случае организм называется гомозиготным по данному гену, во втором - гетерозиготным. Двумя основополагающими принципами - перераспределения генетического материала в поколениях и аллельности генов - и объясняются знаменитые законы Менделя.
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЕЙ
Имея представление о том, что такое аллели, легко объяснить первые законы генетики — три закона Менделя.
Впрочем, самим Грегором Менделем они были выведены во второй половине прошлого века чисто эмпирически на основе результатов проведенных им скрещиваний растений.
Первый закон - так называемый закон единообразия первого поколения при скрещивании чистых форм.
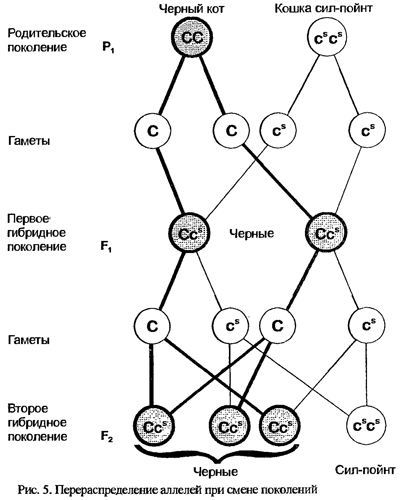
Используем в качестве примера скрещивания таких партнеров: кота и кошку, различающихся по окрасу: кот черный, а кошка - сиамского, или гималайского окраса - сил-пойнт. Причем в обозримом прошлом все их предки имели тот же окрас, то есть оба партнера являются гомозиготными по аллелям интересующего нас гена. Все котята потомки от такого скрещивания будут одинаково черными. Почему?
Сделаем небольшое отступление. Генетическая информация, заложенная в организме, называется генотипом, а ее внешнее проявление - фенотипом. В конкретных случаях часто говорят о генотипе, имея в виду не весь геном, но какой-то отдельный ген или группу генов. Фенотип определяется генотипом прежде всего за счет взаимоотношения аллелей, входящих в состав первого. Каждый обнаруживаемый у животных или человека ген получает свое название, а для обозначения аллелей этого гена используют одну-две начальные буквы его английского наименования (например, White -доминантного белого окраса, чей аллель может быть обозначен как W). Сиамский, или гималайский окрас - это вариант неполного альбинизма (акромеланизма), вызванный мутацией в локусе гена Colour.
Один из аллелей этого гена обеспечивает равномерный полностью развитый окрас, и, как показывают результаты скрещивания, его проявление у потомства преобладает. Такой аллель называется доминантным, и его символ изображается прописной буквой С. Другой аллель - сиамского окраса - является рецессивным (подавляемым) по отношению к С и обозначается строчной буквой "с" с надстрочным символом “s”
Итак, родительские особи были гомозиготные по разным аллелям локуса Colour: кот - СС, а кошка - сs сs(рис.5). Каждый из них может давать только один тип половых клеток (в отношении данного гена, разумеется): сперматозоиды, несущие С, и яйцеклетки, несущие сs.
При объединении этих гамет в новый организм возможен только один вариант генетического набора Ссs то есть все котята будут и гетерозиготные, и единообразны внешне.
Взаимодействие аллелей по типу доминантность - рецессивность наиболее распространено. На уровне механизма действия генов это явление можно объяснить различной активностью образующихся белков (особенно регуляторных): более активные формы будут иметь внешнее выражение, а менее активные или неактивные вовсе не проявятся. Часто аллели, определяющие последние типы белков, называют гипоморфами или аморфами. Если же активность белков примерно равна, можно встретиться с явлением промежуточного наследования. В том же локусе Colour существует еще один аллельный вариант – сb определяющий в гомозиготе другой акромеланистический окрас — бирманский, с незначительным осветлением корпуса и золотистыми глазами. В гетерозиготе с аллелям С (аллелей одного гена у одного организма только пара) сb ведет себя как рецессивный, а вот в сочетании c сs обуславливает промежуточный, тонкинский окрас — более светлый, чем бурма, но темнее сиамского, и специфический зелено-голубой цвет глаз. Тем не менее при скрещивании гомозигот cb cb и cs cs потомство будет опять-таки однородно по окрасу и генетически гетерозиготно - cb cs.