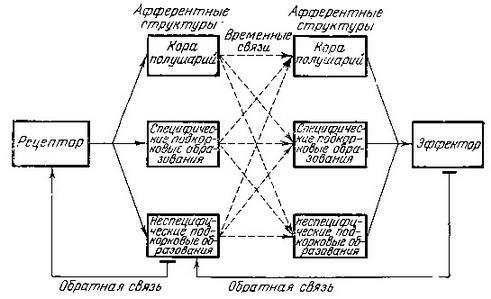
Заслуживает внимания также схема, предложенная несколько лет назад Н. Ю. Беленковым (рис. 31). В ней нашла свое отображение идея о многоканальности условных связей: условные рефлексы могут быть выработаны не только корой большого мозга, но и разными подкорковыми нервными образованиями как самостоятельно, так и при участии коры. По представлениям автора, условные связи могут быть кортикальными, кортико-субкортикальными и субкортикальными, существующими одновременно и образующими сложную систему связей даже при относительно простых формах условных рефлексов.
6. Могут быть отмечены некоторые достижения в изучении явлений торможения в условно-рефлекторной деятельности. В этом плане мы бегло остановимся на трех важных и недостаточно ясных вопросах проблемы кортикального торможения: на механизмах возникновения внутреннего торможения, на локализации этого торможения и на нейрофизиологических основах кортикального торможения.
Рис. 31. Возможные пути замыкания условного рефлекса и общий принцип его структуры (по Беленкову)
Как было отмечено, несмотря на выдающиеся достижения, сделанные Павловым в экспериментальном и теоретическом изучении форм, механизмов возникновения и роли торможения в деятельности большого мозга, он очень осторожно, с постоянными оговорками и сомнениями высказывался по тем или иным вопросам кортикального торможения. Ученый сам мотивировал это ограниченностью накопленного фактического материала, недостатком точных знаний об интимных нейрофизиологических обновах, крайней сложностью проблемы, трудностью ее экспериментальной разработки и особенно теоретического освещения и другими обстоятельствами, побудившими его назвать саму проблему «проклятой». Весьма показательно в этом отношении резюмирующее высказывание Павлова, сделанное в одной из своих знаменитых «Лекций», посвященной специально проблеме торможения: «Вот почему,— говорил он,— я пока отказываюсь высказываться за ту или другую из существующих теорий торможения или выставлять новую. Но мы пользуемся временными предположениями для систематизирования нашего фактического материала и проектирования новых опытов» [1 И. П. Павлов. Полн. собр. трудов, т. IV, стр. 320—321.].
Допущение Павлова о локализации внутреннего торможения в клетках кортикального пункта условного раздражителя было одним из наиболее слабых звеньев в системе его взглядов на торможение. Если отдельные факты, полученные в его лаборатории, и могли служить основанием для этого допущения (скажем, описанные выше данные о порядке и продолжительности иррадиации и концентрации условного торможения), то ряд других общеизвестных и достоверных фактов, накопленных им и его сотрудниками, явно противоречил такому допущению. Но ученый почему-то должным образом не учитывал их. К ним относится, в частности, установленный Г. В. Миштовтом и другими сотрудниками давний факт о том, что тормозной условный раздражитель вызывает у животного активнук> реакцию, по характеру антагонистическую реакции на положительный условный раздражитель. Можно назвать также давний факт, полученный Л. Н. Воскресенским, В. М. Архангельским и другими сотрудциками лаборатории, относительно того, что секреторный и двигательный компоненты пищевого условного рефлекса при угасании, дифференцировке и прочих разновидностях внутреннего торможения тормозятся не одновременно: по одному показателю, рефлекс затормаживается, а по другому еще четко проявляется. Совершенно очевидно, что если бы при этом условное торможение локализовалось в клетках кортикального пункта условного раздражителя, то названные явления не могли бы иметь места: действие тормозного раздражителя не вызвало бы каких-либо реакций, а двигательный и секреторный компоненты условного рефлекса должны были бы иметь синхронную динамику торможения.
И действительно, в дальнейшем многими учениками и последователями Павлова в новых опытах было показано, что внутреннее торможение возникает и локализуется не в нервных элементах кортикального пункта условного раздражителя, как считал ученый, а в одном из следующих звеньев дуги условного рефлекса (И. Я. Перельцвейг, Н. А. Кашерининова, Д. С. Фурсиков, П. К. Анохин, П. С. Купалов, Э. А. Асратян). Например, в нашей лаборатории установили, что если на один и тот же раздражитель вырабатываются два разного рода условных рефлекса, скажем пищевой и электрооборонительный (так называемые бинарные условные рефлексы), то при угасании одного из них второй остается в полной сохранности. Более того, электрофизиологическими экспериментами Л. С. Чистович, Г. Ю. Кратина, У. Г. Гасанова и др. показано, что развитие условного торможения влечет за собой даже повышение возбудимости кортикального пункта раздражителя заторможенного рефлекса, по меньшей мере в начальных фазах развития торможения. Интересны в этом отношении также факты лабораторий Г. В. Гершуни, М. М. Кольцовой, Н. И. Касаткина и др. о восприятиях детьми и взрослыми тормозных условных раздражителей так же, как и положительных.
Следует сказать, что в настоящее время почти общепринято, что условное торможение возникает и локализуется не в кортикальных структурах условного раздражителя, как считал Павлов. Однако нерешенным еще остается вопрос: в каком же из следующих звеньев дуги условного рефлекса это происходит? Мнения исследователей этого вопроса сильно расходятся. Одни считают таким местом кортикальные или даже субкортикальные структуры подкрепляющего безусловного раздражителя (И. Я. Перельцвейг, Н. А. Кашерининова, П. К. Анохин, Б. И. Ходоров и др.), другие — мозговые структуры и условного и безусловного раздражителей (П. С. Купалов и др.), третьи разделяют развиваемую нами точку зрения о том, что это торможение возникает и локализуется в условной связи (И. С. Бериташвили, Л. Г. Воронин, Ф. П. Майоров и др.). Ввиду узкоспециального характера этого вопроса нет необходимости приводить здесь факты и доводы в пользу каждой из упомянутых выше точек зрения. Отметим только, что в его окончательном разрешении весьма важную роль призваны играть новейшие микро-электрофизиологические методики.
Существенно расходятся мнения исследователей также но вопросу о механизмах возникновения внутреннего торможения. Точка зрения Павлова была уже кратко изложена в одной из предыдущих глав. Она сводится к допущению, что при отмене подкрепления условного рефлекса хрупкие по природе нервные клетки кортикального пункта условного раздражителя лишаются индукционного торможения как профилактического фактора (которое обычно в них развивается под влиянием центра безусловного рефлекса, сильно возбужденного подкрепляющим раздражителем), остаются возбужденными продолжительное время и истощаются. При повторных неподкреплениях условного рефлекса это истощение углубляется все больше и больше, в клетках происходит процесс функционального разрушения и на этом фоне возникает и постепенно усиливаемся торможение как охранительный и восстановительный фактор, как физиологическая мера самозащиты; это торможение в последующем становится внутренним торможением с весьма важной координационной ролью.
Этой точки зрения придерживаются многие из последователей великого физиолога и в настоящее время, причем некоторые из них (В. К. Федоров, Э. А. Асратян и др.) сочли возможным на основании нового фактического материала внести в исходную форму концепции учителя существенные коррекции, модифицировать ее в том или ином пункте. Например, в нашем варианте допускается, что при отмене подкрепления первыми истощаются не кортикальные клетки условного раздражителя и не в них изначально возникает охранительно-восстановительное торможение, как считал Павлов, а нервные структуры самой условной связи, т. е. среднего звена дуги условного рефлекса, и что торможение сначала появляется и локализуется именно в них.
Следует отметить, что и в новых модификациях исходной концепции Павлова вопрос об интимном характере процесса преобразования охранительно-восстановительного торможения во внутреннее торможение остается по существу нерешенным.