Так, диффузная активация структур центральной нервной системы при стрессе обусловлена их связями с ретикулярной (сетчатой) формацией ствола головного мозга. Ее можно рассматривать как систему первичной структуризации энергии, выделяемой в процессе обработки информации, и, тем самым, – как механизм предупреждения дальнейшего роста энтропии. Возможно, это наиболее древняя функция нервной системы, так как в эволюции, впервые возникнув у кишечнополостных (Hydroidea), она имеет сетчатую структуру, подобно ретикулярной формации мозга позвоночных. Модельные эксперименты, проведенные на гидрах с фармакологически выключенной нервной системой, показали, что функции других систем сохраняются относительно постоянными, но укорачивается продолжительность жизни животных, что может быть следствием ускоренного роста уровня обобщенной энтропии.
В эволюции нервной системы прослеживается усиление роли и другого фактора снижения энтропии, – развития тормозных систем и увеличения разнообразия тормозных медиаторов, выделяемых нейронами и глиоцитами. На активную роль торможения в организации живого организма как биосистемы, особенно в связи с концентрацией внимания, формированием доминанты, впервые указал А. А. Ухтомский (Ухтомский, 1966).
Еще одним механизмом снижения обобщенной энтропии в живых организмах является асимметрия (Чернышева, 2003). Хотя структурно-функциональная асимметрия является одним из характерных свойств многоклеточных, но ее энергетическая «выгодность» по сравнению с симметрией была показана и на атомарном уровне (Мøller e.a., 2002): при асимметричном делении лучом лазера ядер гелия и фермия энергетический порог ниже, а суммарная кинетическая энергия осколков ядер выше, чем при симметричном.
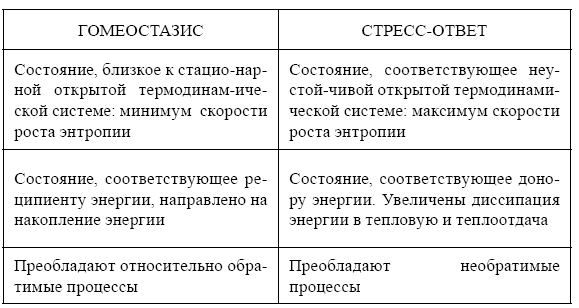
Таблица 1.
Сравнительная термодинамическая характеристика функциональных состояний гомеостазиса и стресс-ответа организма (по: Чернышева, Ноздрачев, 2006)
В живых организмах асимметрия как феномен обладает уникальными свойствами. С одной стороны, она повышает неустойчивость организма и, следовательно, его энергетический потенциал, способствуя увеличению скорости обмена веществ и уровня энтропии, росту возбудимости и, сопряженно, – увеличению сенсорной чувствительности и объема воспринимаемой информации. Последнее, с другой стороны, определяет вклад асимметрии в сдерживание роста обобщенной энтропии. Этому способствуют и проявления морфо-функциональной асимметрии, описанные для парных структур. Например, меж-полушарная асимметрия головного мозга животных и человека (Сhernysheva, 2006; Nikolaeva, Leutin, 2011) заключается в различии не только ряда функций двух полушарий, но и уровня обмена веществ и обобщенной энтропии. Так, правое полушарие обладает более высоким уровнем обмена веществ и энергии, обобщенной энтропии, тогда как левое – более низким (Gur et al., 2002; Andrew, 2002; Чернышева, 2003). Взаимосодействие парных структур направлено на снижение энергозатрат как «цены адаптации».
Взаимосвязь асимметрии с регуляцией уровня энергообмена организма подчеркивает усиление ее при стрессе. О важности феномена свидетельствует также эволюция проявлений морфо-функциональной асимметрии на всех уровнях организации живого. Примером может служить быстрый переход в эволюции плана тела животных от пятилучевой симметрии к трехосевой асимметрии, что отражает процесс адаптации плана тела живых организмов к трехмерному окружающему пространству.
Итак, перечислим кратко свойства живых организмов, позволяющие снижать рост обобщенной энтропии вопреки второму началу термодинамики. Среди них: сопряжение обратимых и необратимых процессов, структурная и функциональная организация потоков энергии, эндогенные источники энергии и информации, память, концентрация внимания, доминанта, торможение, асимметрия, способность регулировать гомеостазис и степень «открытости» организма как неустойчивой термодинамической системы во взаимодействии с окружающей средой. Эти свойства позволяют поддерживать достаточно низкую скорость роста энтропии, а также быть относительно независимыми от окружающей среды. Можно предположить, что термодинамическая «пластичность» и разнообразие путей «обхода» запрета второго начала термодинамики живыми организмами являются весомыми факторами, определяющими специфику живого (Чернышева, Ноздрачев, 2006). Кроме того, процессы жизнеобеспечения, связанные с делением, ростом и дифференцировкой клеток, метаморфозами и регенерацией, движением и поведением, не приводят к «тепловой смерти», но сохраняются в течение жизни и могут передаваться генетически благодаря названным «антиэнтропийным механизмам» (термин Ю. А. Романова, 2000).
Общеизвестно, что живые организмы как открытые термо-динамические системы обмениваются с окружающей средой материей, энергией, информацией и, добавим, временем. Последнее согласуется с тезисом о существовании времени только для открытых систем (Левич, 2013). Для успешности такого обмена необходимым условием является способность биосистемы создавать материю, генерировать энергию, информацию и время. Это подтверждают способность к образованию молекул веществ в процессах синтеза, метаболизма нутриентов поглощаемой пищи и катаболизма синтезированных веществ, а также выделение энергии в реакциях метаболизма, дефосфорилирования макроэргов (АТФ, ГТФ, КФ и др.) и других молекул или же их депротонирования и т. д. В частности, известно, что в процессах генеза и процессинга информации в нервной системе усиление активности Na,K-ATФазы клеточной мембраны нейрона на фазе следовой гиперполяризации потенциала действия приводит к восстановлению асимметрии концентрации ионов Na+ и K+ и потенциала покоя мембраны, а реаптейк транспортерами молекул нейромедиатора на уровне пресинаптической мембраны осуществляется на градиенте Н+ или Na+. Известно, что процессы сокращения и расслабления скелетных мышц при поддержании позы или двигательной активности также осуществляются при участии Na,K-ATPазы и Са,Mg-ATPазы, способных присоединять молекулы АТФ, дефосфорилировать их, а выделившуюся энергию частично использовать на перенос ионов через мембраны против градиента их концентраций. Каждый из этих процессов характеризуют временные параметры (латентность, длительность, скорость), что позволяет говорить о них как о временных процессах.
Постулируем взаимосвязь биологического времени с информацией, метаболизмом и энергией. Для формализации такой взаимосвязи рассмотрим особенности взаимодействия информации и времени, а также метаболизма и времени.
1.2. Информация и биологическое время
Известный тезис Аристотеля «Время является мерой движения (изменения)» (Аристотель, 1937) применительно к живым организмам может быть переформулирован как «время есть мера изменения информации» В пользу этого тезиса свидетельствует ряд исследований, в частности, работа R.E. Hicks и соавторов (Hicks et al., 1976), в которой авторы рассматривают проспективные и ретроспективные суждения о времени как функцию от объема полученной информации. О схожей закономерности, связывающей время и информацию в биосистемах писал М. И. Сетров (1974).
Проанализируем в этом аспекте два основные определения: информация как сообщение/сигнал о чем-либо и информация как негэнтропия (Шредингер, 2002; Бриллюэн, 2006). Другие определения условно можно считать по смыслу близкими первому или второму из них, дополняющими характеристику свойств/функций информации. Многочисленные данные из различных областей биологии свидетельствуют о справедливости для биосистем обоих определений, а также об одновременном взаимосвязанном генезе информации и эндогенного времени на разных структурных уровнях организма (Чернышева, 2011). Рассмотрим эти положения более конкретно.
1.2.1. Информация как сигнал/сообщение
Известно, что рецепторы живых организмов как специфические сенсорные структуры воспринимают и усиливают экзо- или эндогенные воздействия определенной энергетической природы, а также передают сигнал о них далее, в нервные центры. Так, зрительные рецепторы активируются энергией света, тогда как обонятельные, вкусовые и хеморецепторы сосудов и внутренних органов – энергией химических взаимодействий рецепторов с одорантами, нутриентами или продуктами обмена веществ. Разнообразные рецепторы опорно-двигательной системы, рецепторы прикосновения и давления кожи, барорецепторы сосудов, а также слуховые и гравитационные рецепторы воспринимают воздействия факторов, сопряженных с механической энергией. В рецепторных нервных окончаниях воздействие определенной энергетической природы приводит к возникновению рецепторного и, затем, генераторного потенциала, что отражает генез информации о воздействии. Ее внутриклеточным кодом являются кальциевые спайки, распространяющиеся по внутренним структурам дендрита (Sjöström et al., 2008, и др.) к соме и, затем, к аксону сенсорного нейрона. Возникающие под его влиянием в начальном сегменте аксона потенциалы действия (спайки или импульсы) отражают усиление, кодирование и передачу информации другим клеткам. Потенциал действия возникает как изменение мембранного потенциала (электрического сигнала) в результате трансмембранного движения ионов натрия и/или кальция, а также калия и хлора через соответствующие ионные каналы. Для передачи информационного сигнала следующему нейрону или иной клетке-эффектору путем выделения определенного химического медиатора важно, чтобы последовательность потенциалов действия включала более двух спайков, следующих с определенной частотой. Экспериментально доказано, что при разной частоте импульсов аксон может выделять разные комплексы медиаторов и ко-медиаторов. Это свидетельствует об электро-хемо-частотной (или – временнóй) природе первичного кода информации о воздействии. Таким образом, воздействия разной энергетической природы описываются универсальным электро-хемо-временным «языком». При этом генез информации взаимосвязан с возникновением не только временнóго компонента кода, но и совокупности процессов, отраженных в генерации потенциалов, кодировании, усилении и передаче сигнала от мембранных структур к внутриклеточным, а также от клетки к клетке. Каждый из них обладает набором темпоральных параметров (латентностью, скоростью, длительностью), что позволяет называть эти процессы временны́ми и считать их компонентами эндогенного (биологического) времени, генерируемого в структурах организма.