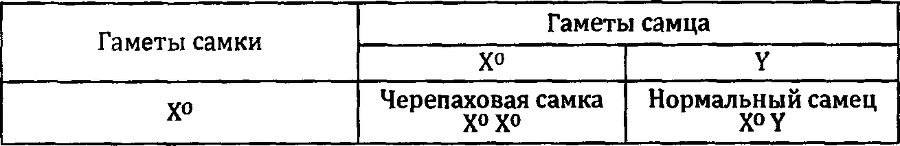Итак, мы с вами переоткрыли на кошке законы Менделя. Попытаемся суммировать наши переоткрытия.
Наследственность дискретна, и вклад каждого родителя одинаков. Гены, полученные от одного родителя, не загрязняются, не разбавляются генами другого родителя. Они остаются неизменными безотносительно к тому, в компании с какими генами они побывали. Так, черный кот, полученный от двух серых гетерозиготных родителей, не менее черен,,чем тот, что получен от скрещивания двух черных гомозигот.
Каждый ген может существовать в двух или более альтернативных формах, которые называются аллелями. Так, у кошки ген, контролирующий длину шерсти, представлен в двух формах: аллеля L, дающего короткую шерсть, и / — длинную. Известны три аллеля гена, контролирующего рисунок: Та — абиссинский тэбби, Т — тигровый тэбби и tP — мраморный, или пятнистый тэбби.
Один аллель может полностью или частично доминировать над другим. Доминантные гены определяют фенотип как в гетеро- так и в гомозиготном состоянии. Рецессивные же — только в гомозиготном. Аллель А — агути-окраска — проявляется и при одной, и при двух его дозах в организме. Аллель а — черная, не- агути окраска, проявляется только в том случае, если он присутствует в двух дозах. Проявление аллеля S зависит от его дозы. В двойной дозе он дает большую белую пегость, а в одинарной — пегость средних размеров.
Из двух аллелей родителя потомок получает только один. Другой аллель данного гена он приобретает от другого родителя. Благодаря этому, потомки от скрещивания двух гомозигот всегда одинаковы. Однако в результате скрещивания этих потомков друг с другом рождается разнообразное потомство. Все потомки от скрещивания серого кота с черной кошкой — серы. Но при скрещивании этих потомков первого поколения друг с другом получаются как серые, так и черные потомки.
Если мы следим за расщеплением не по одному, а по двум или более признакам, то легко обнаруживаем, что расщепление по одному признаку не зависит от расщепления по другому. Длина и цвет шерсти распределяются в потомстве безотносительно тому, как они проявлялись у родителей — вместе или отдельно.
В общей форме законы Менделя справедливы для всех диплоидных организмов, к которым относимся и мы, и наши кошки. В основе законов Менделя лежат правила поведения хромосом в мейозе. Независимое расщепление обусловлено тем, что разные гомологи расходятся по гаметам независимо друг от друга.
Вскоре после переоткрытия законов Менделя стали обнаруживаться многочисленные исключения из них. Мы их сейчас кратко рассмотрим. Эти исключения не менее важны для понимания механизмов наследования признаков, чем сами законы.
Начнем с исключений из первого закона — закона единообразия гибридов первого поколения. Скрестим кота, гомозиготного по аллелю белой пегости 5 с кошкой, лишенной белых пятен. Все гибриды первого поколения будут иметь белое пятно и в этом отношении будут единообразны. Но размер пегости у гетерозигот будет варьировать от кошки к кошке. У одних белое пятно будет побольше, у других — поменьше. Такие признаки с изменчивым проявлением генетики называют признаками с варьирующей экспрессивностью. Под экспрессивностью здесь мы понимаем степень проявления признака.
Вариации размеров пятна могут быть обусловлены случайными отклонениями в ходе нормального развития, которые могут повлиять на подвижность меланобластов, на плотность тканей, сквозь которые меланобласты вынуждены мигрировать, на скорость созревания волосяных фолликулов и т. п. На эти же параметры могут оказывать влияние и другие гены, не имеющие прямого отношения к гену S.
Влияние внешней среды на развитие признака может быть очень существенным. Вы знаете, что у гомозигот по аллелю сиамского альбинизма пигментация подавлена на теле и довольно выражена на ушах, носу, лапах и хвосте. Русские исследователи Ильины еще в 1930-х годах показали, что, меняя температуру, при которой протекает детство сиамского котенка, можно управлять степенью его пигментации. Она усиливается при низкой температуре и ослабляется при высокой.
Некоторые признаки могут вообще не проявляться, несмотря на наличие в организме контролирующего их аллеля. О таких признаках с неполным проявлением (пенетрантностью) мы уже говорили в предыдущей главе. Понятно, что наследование таких признаков будет отклоняться от законов Менделя.
Серьезные нарушения первого закона мы обнаруживаем, наблюдая за наследованием признаков, сцепленных с полом, то есть таких признаков, которые контролируются генами, находящимися в половых хромосомах. Мы уже не раз рассматривали аллель рыжей окраски. Вернемся к нему еще.
Скрестим рыжего кота с серой кошкой. По первому закону Менделя все их потомство должно быть одинаковым. На самом же деле в первом поколении мы получаем два класса потомков: все самцы серые, все самки — черепаховые. Скрестим рыжую кошку с серым котом. В потомстве от этого скрещивания опять будут разные животные: рыжие самцы и черепаховые самки. Причины черепаховости мы уже разбирали. А причины нарушения первого закона Менделя для этого случая иллюстрирует решетка Пеннета.
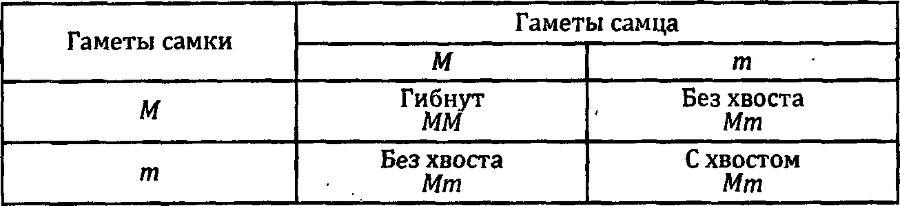
Нарушения второго закона Менделя — закона расщепления — часто наблюдаются при скрещивании гетерозигот по летальным или сильно снижающим жизнеспособность генам. Так, если мы скрещиваем двух бесхвостых кошек, гетерозиготных по аллелю мэнской бесхвостости М, то получается расщепление 2:1 (2 бесхвостых и 1 хвостатый) вместо менделевского 3:1. Причина нарушения в том, что гомозиготы ММ гибнут до рождения.
Именно поэтому невозможно получить чистую, нерасщепляющуюся породу мэнских бесхвостых кошек: при скрещивании бесхвостых всегда будут рождаться и хвостатые потомки.
Очень интересны нарушения третьего закона Менделя — закона независимого расщепления признаков при скрещивании особей, различающихся не по одному, а по двум или большему числу признаков. Причины и следствия этих нарушений могут
быть самыми разными. Поговорим о некоторых из них.
Одной из причин зависимого расщепления двух признаков может быть сцепление генов. Мы говорили о том, что гены не болтаются в клетке по отдельности, а сцеплены друг с другом в составе хромосом. Если 30 000 генов разделить на 19 пар хромосом, то на каждую хромосому в среднем придется более чем по 1000 генов. До сих пор мы рассматривали гены, которые входят в состав разных, негомологичных хромосом. Естественно, что такие гены расходятся по гаметам независимо друг от друга. Если же мы возьмем гены, находящиеся в одной хромосоме, то обнаружим, что они расходятся согласованно. Если бы ген черной окраски был тесно сцеплен с геном-ослабителем, то во втором поколении от описанного выше скрещивания голубого кота с серой полосатой кошкой мы получили бы расщепление не на 16 генотипических классов, а только на 4. Вместо четырех типов гамет образовалась бы только два AD и ad.
Я не случайно употребил здесь выражение «тесно сцеплены». Только в том случае, если два гена находятся в хромосоме очень близко, они почти всегда будут расходиться по гаметам вместе. Если же один из них расположен от другого на некотором расстоянии, то возникает возможность расхождения аллелей этих двух генов в разные гаметы.
Мы говорили с вами о том, что в ходе подготовки к первому делению мейоза гомологичные хромосомы способны обмениваться отдельными участками, рекомбинировать гены. Чем дальше друг от друга расположены два гена в хромосоме, тем более вероятен обмен, тем больше характер расщепления по контролируемым ими признакам будет приближаться к тому, что предписан третьим законом Менделя.
Таким образом, по степени нарушения этого закона каждой конкретной парой генов мы можем судить о расстоянии между ними на хромосоме. На этой закономерности основан рекомбинационный принцип создания генетических карт. С использованием этого метода было установлено, что гены белой пегости 5 и полидактилии Pd находятся в одной хромосоме на расстоянии, равном 35% ее длины.