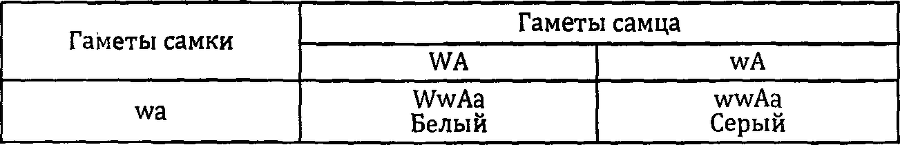Когда мы говорим о третьем законе Менделя, следует помнить, что он справедлив (с указанными выше ограничениями! для тех случаев, когда признаки контролируются разными генами. Например, цвет и длина шерсти зависят от разных генов и наследуются независимо. Цвет шерсти и острота слуха — безусловно, различные признаки. Они контролируются разными генами, но нарушения и того, и другого признака могут вызываться одной мутацией.
Яркий пример такого рода множественного, или плейотропного действия гена — проявление мутации доминантной белой окраски W. Гомо- и гетерозиготные носители этого аллеля белые, но, кроме того, они часто имеют голубые глаза и ослабленный слух. Здесь не следует ожидать независимого расщепления по этим трем признакам именно потому, что нарушения в каждом из них вызываются мутацией одного гена.
Есть, однако, много примеров противоположного свойства: когда за развитие одной системы отвечают несколько генов. Вы знаете, что на окраску шерсти влияет и скорость миграции меланобластов, и интенсивность синтеза пигментов в меланоцитах, и распределение пигментных гранул в волосе. Результирующий признак — окраска шерсти — зависит от согласованного действия генов, отвечающих за каждый из этих процессов. Попробуем разобраться с характером этого взаимодействия.
Один мой знакомый скрестил своего белого кота с черной кошкой. В потомстве от этого скрещивания он получил двух белых и двух серых котят. Этот результат его так удивил («Откуда взялись серые?!»), что он устроил скандал хозяйке черной кошки. Хозяйка резонно возразила ему, что его возмущения были бы понятны, если бы котята были не похожи на отца, но они-то не похожи на мать! После жарких дебатов о том, кто виноват, они перешли к вопросу, что делать, и решили обратиться ко мне. Это был первый разумный поступок участников этой истории.
Выслушав их внимательно, я надул щеки, пошевелил усами и сказал:
— Что такое генетика, друзья мои? Генетика — это наука, которая объясняет, почему ребенок похож на отца, если он на него действительно похож, и почему не похож, если уж так получилось. Не надо оваций. Это не моя мудрость. Это из «непричесанных мыслей» Ежи Леца. Моя же мудрость заключается в том, что я вам сейчас объясню, почему родились котята, не похожие ни на отца, ни на мать: серые от белого и черной. Все дело в доминантном эпистазе.
Нельзя сказать, чтобы это мое последнее заявление очень успокоило моих слушателей. Я вообще заметил, что генетические термины действуют успокаивающе только на генетиков. Если вы говорите генетику, что абсурдное расщепление, которое он получил, обусловлено неполной пенетрантностью, то генетик сразу примиряется с этим расщеплением, хотя никто толком не знает, в чем причины неполной пенетрантности и что вообще за этим словом стоит. Но на нормальных людей генетические термины действуют, как на котов котовник кошачий: они приходят в ярость.
Поэтому я вынужден был пуститься в более подробные объяснения. Я указал им на то, что серая окраска доминирует над черной, а отсутствие окраски — над ее наличием: А над a, Wнад w. Исходя из этого и из результатов скрещивания легко установить генотипы родителей. Генотип отца WwAA, г,енотип матери wwaa. Построим решетку Пеннета.
Расщепление по фенотипам идеально соответствует тому, что было получено в результате скрещивания белого кота с черной кошкой. А если уж говорить человеческим языком, то этот кот был бы серым, кабы его шерсть была окрашена. Но поскольку она не имеет окраски, то ничего о том, какой аллель гена агути он содержит, сказать нельзя. Ответ дает анализирующее скрещивание. Под покровом белой окраски наш кот мог нести и ген рыжей окраски, и ген белого пятна. Проявление этих генов было подавлено эпистатическим действием аллеля белой окраски. Эпистазом мы называем подавление одним геном проявления другого гена, неаллельного первому.
Если есть доминантный эпистаз, то должен быть и рецессивный. У кошек это явление можно проиллюстрировать на примере аллеля с° — голубоглазый альбинос. Гомозиготы по этому аллелю совершенно белые со светло-голубыми глазами.^
Такого гомозиготного самца скрестили с черной самкой. В первом поколении от этого скрещивания все котята были серыми. Этот результат показывает, что наш белый кот гомозиготен по аллелю А. Теперь, когда мы знаем генотипы родителей, можно записать генотипы потомков первого поколения. Все они имеют генотип АаСса.
Скрестим их друг с другом. В потомстве мы получим девять серых, трех черных и четырех белых голубоглазых потомков. Решетка Пеннета дает нам объяснение, почему расщепление в данном случае именно такое, а не иное.
Рецессивный эпистаз
Вообще генетику гораздо интереснее те случаи, когда ребенок не похож на родителей, чем те, когда он похож. В разобранном нами случае часть котят была не похожа ни на отца, ни на мать. Но бывают интересные ситуации, когда ни один из котят не похож ни на одного из родителей. Именно такую мы сейчас проанализи руем.
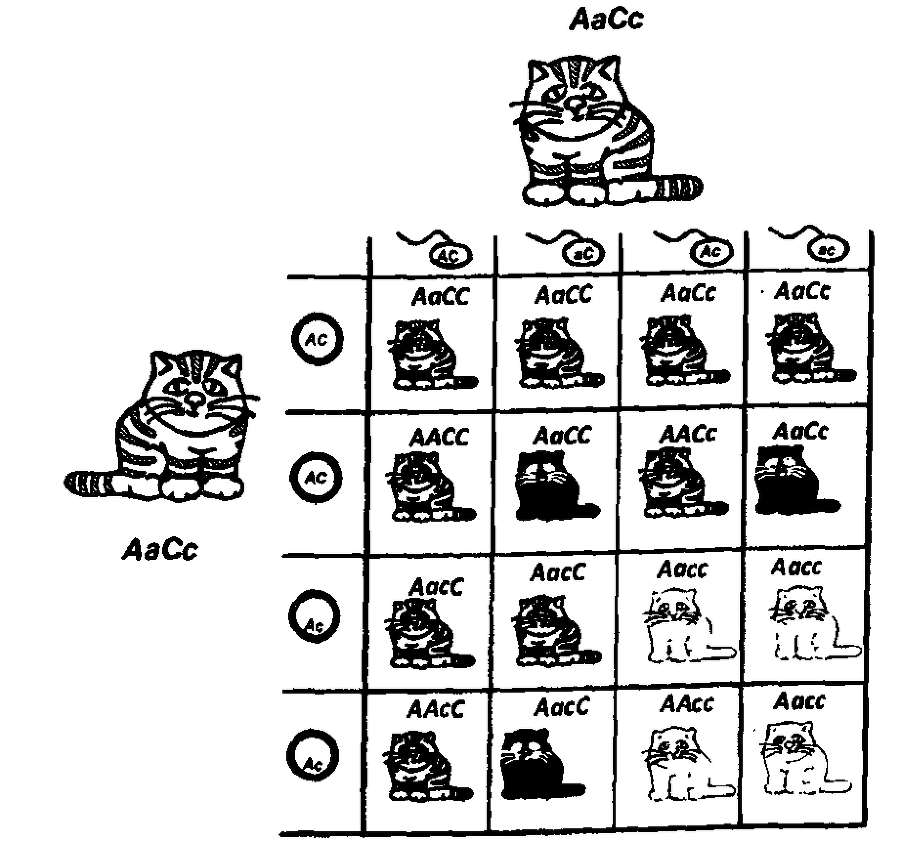
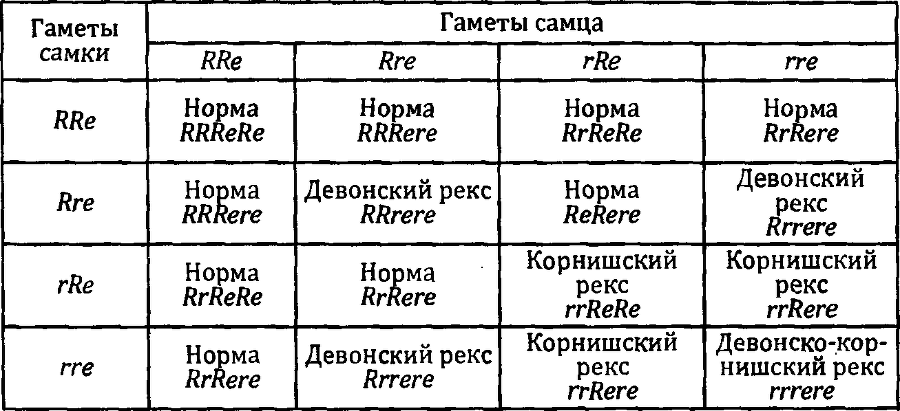
В предыдущей главе я говорил о том, что обнаружено несколько мутаций, дающих в гомозиготе рексовую текстуру шерсти. Корнишский реке приводит к полной элиминации остевых волос, а девонский — к частичной. Но в целом гомозиготы по каждой из этих мутаций очень похожи друг на друга. Что будет, если скрестить чистопородного девонского рекса с чистопородным же рексом, но корнишским?
Все потомки от такого, скрещивания будут с нормальной шерстью. Почему? Потому, что рексовость вызвана мутациями в разных генах. Обе мутации рецессивны. При скрещивании мы получаем гетерозигот по каждому из генов, и это приводит к восстановлению нормального внешнего вида.
Если мы скрестим этих животных первого поколения с нормальной шерстью, друг с другом, то получим расщепление, которое мы уже разбирали, говоря о третьем законе Менделя: девять нормальных, три корнишских рекса, три девонских рекса и один, имеющий признаки обоих рексов сразу.
По-видимому, каждый из этих двух генов отвечает за разные этапы формирования волоса. Однако нарушения, происходящие на любом из этапов, ведут к одному результату.
В первой главе мы упоминали о том, что некоторые гены присутствуют в геноме в нескольких экземплярах. Каждый из этих генов может вносить определенный вклад в формирование признака, усиливая или ослабляя его проявление. Результат в таком случае будет зависеть от числа усиливающих и ослабляющих аллелей в геноме каждого животного. Как правило, под контролем таких многократно повторенных генов — полигенов — находятся количественные признаки.
У кошек к количественным признакам, которые могут заинтересовать кошковладельцев, относятся длина хвоста, длина тела, ширина морды и другие. Рассмотрим, насколько применимы к таким признакам законы Менделя.
Допустим, что на длину хвоста влияют гены А, В и С. Пусть каждый из них определяет 10 сантиметров хвоста. Допустим также, что существуют мутантные аллели, каждый из которых в гомозиготе ведет к укорочению хвоста на 10, а в гетерозиготе — на 5 сантиметров. При этих допущениях положим, что кот с генотипом ААВВСС имеет хвост длиной 30 сантиметров, а кошка aabbcc вовсе лишена хвоста. Скрестив их друг с другом, мы получим потомков первого поколения с хвостами средней длины 15 сантиметров и выполним тем самым первый закон Менделя — закон единообразия первого поколения.
Проведя скрещивания гибридов первого поколения друг с другом, мы получим биномиальное распределение потомков второго поколения по длине хвоста. Больше всего будет потомков со средней длиной хвоста, а меньше всего тех, которые похожи на своих бабушку и дедушку.
Если мы отложим по оси абсцисс длину хвоста, а по оси ординат — число особей, обладающих хвостом данной длины, то распределение по длине хвоста у гибридов второго поколения от скрещивания длиннохвостого кота с бесхвостой кошкой будет иметь вид типичного биномиального распределения. Чем больше генов участвует в контроле интересующего нас признака, тем более плавным будет это распределение.