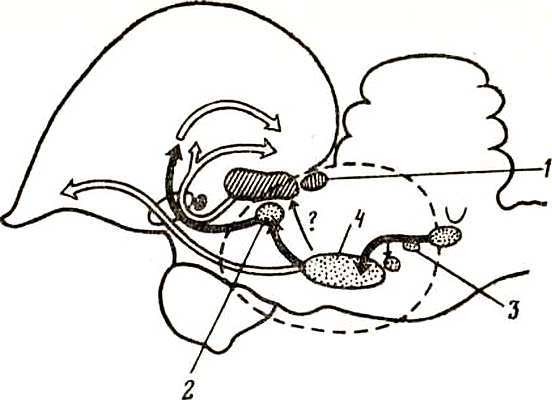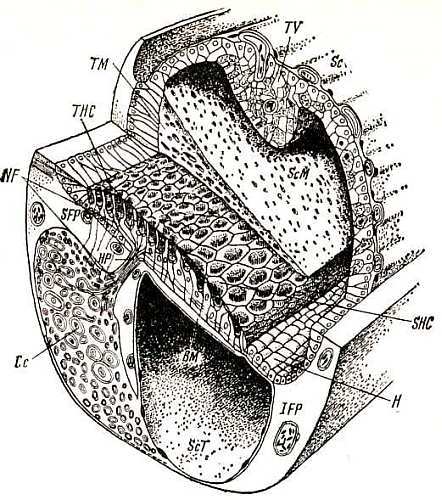
Рис. 7. Микроструктуры улитки птицы (Takasaka, Smith, 1971) Рецепторный эпителий, состоящий из чувствительных и опорных клеток, располагается на базилярной мембране (ВМ). Скошенные (кувшиновидные) волосковые клетки (SHC) лежат на свободной базилярной мембране, высокие волосковые клетки (THC) — на переднем хряще (SFP). Текториальная мембрана (ТМ), часть которой удалена, чтобы показать поверхность рецепторного эпителия, прикрепляется к передним гомогенным клеткам и покрывает поверхность рецепторного эпителия. На задней зоне базилярной мембраны расположены задние гиалиновые клетки (Н), от рецепторного эпителия они отделены кубическими клетками. Сосудистая обкладка (TV), прикрепляясь к переднему и заднему (IFP) хрящам, отделяет кохлеарный проток (ScM) от вестибулярной лестницы (ScV), которая на рисунке (вверху справа) открыта. Показаны периферические отростки нейронов кохлеарного ганглия (Сс), проникающие в рецепторный эпителий через habenula perforata (HP). Под базилярной мембраной — тимпанальная лестница (ScT)
Базилярная мембрана птиц подложена широкими пластинами хрящей, оставляющими свободной только самую центральную ее часть в виде узкой продольной полоски. При этом длина базилярной мембраны и длина рецепторного эпителия в улитке серой вороны значительно превышают таковые голубя (соответственно 4,4 и 3,3 мм). Конец улитки заполнен особым образованием — лагеной. Лагена представлена группой волосковых клеток, прикрытых студенистой макулой с кристаллическими образованиями. Внешне это напоминает рецепторы вестибулярного аппарата и эту функцию лагене долго приписывали. В последние годы, однако, выяснилось, что нервные волокна, идущие от лагены, направляются далее в слуховые ядра мозга (как и в вестибулярные), и, следовательно, лагена может воспринимать звуки. У млекопитающих улитка не имеет лагены вовсе.
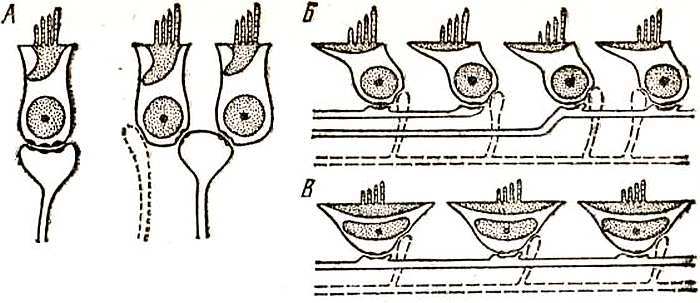
Рис. 8. Типы волосковых клеток и их иннервации у птиц (Голубева, Ямалова, 1980) А — иннервация высоких волосковых клеток, В — скошенных клеток, В — низких волосковых клеток. Сплошной линией обозначены афферентные нервные волокна, пунктирной — эфферентные
Один из вопросов, который стоит в связи с изучением улитки птиц, заключается в следующем. Что она представляет собой как звено слуховой системы? Нечто вроде «проходного двора», пропускающего наверх все, что к ней приходит, или она принимает участие в обработке звука, в анализе слуховой информации?
Например, одной из важнейших функций улитки млекопитающих является ее участие в анализе частоты звука. Базилярная мембрана узка в начальной части улитки и широка в конечной. Это обеспечивает настройку на разные частоты различных участков мембраны… Звук в виде механических колебаний передается слуховыми косточками на жидкость внутреннего уха и через нее на базилярную мембрану. На мембране возникает бегущая изгибная волна, распространяющаяся от основания улитки к ее вершине. В определенном участке улитки, настроенном на данную частоту, волна достигает максимума, деформируя в этом участке покровную мембрану и возбуждая волосковые клетки. Возбуждение последних через отростки нейронов передается на кохлеарные ядра продолговатого мозга и направляется вверх к слуховой коре. Такова сильно упрощенная схема. Но можно ли ее перенести на улитку птиц? Только с очень большой осторожностью. Базилярная мембрана здесь много короче и относительно шире, однако большая ее часть подостлана хрящом и не свободна. Рецепторные клетки не имеют строго продольной ориентации. Большая рыхлая сосудистая покрышка имеется у птиц и отсутствует у млекопитающих. Различия, как мы видим, достаточно велики.
Вместе с тем имеются данные, доказывающие, что улитка птиц также принимает участие в анализе частоты.
Советский ученый О. III. Гогниашвили показал, что действие звука разных частот возбуждает рецепторы строго определенных участков базилярной мембраны, что особенно интересно, — продольных, а не поперечных рядов. Как будто бы волна распространяется не вдоль мембраны, а поперек ее. Эти работы были выполнены с помощью гистохимических методик. А профессор И. Шварцкопф из Рурского университета и профессор М. Конинга из Принстонского показали, что хотя слуховые нейроны продолговатого мозга настроены на определенные частоты, каждый на свою, но таким образом, чтобы охватить достаточно широкий диапазон воспринимаемых частот.
У птиц, особенно нуждающихся в тонком различении частот, улитка оказалась длиннее, число нейронов в нерве и слуховых ядрах больше. Сотрудница Московского университета Л. И. Барсова обратила внимание на особое расположение нейронов некоторых ядер, непосредственно связанных с улиткой. Именно в этих ядрах скандинавский гистолог Р. Бурд нашел отчетливые проекции строго определенных участков улитки. А работы Л. И. Барсовой показали, что нейроны в таких ядрах у видов, особенно нуждающихся в тонком частотном анализе (совы), располагаются в виде плотных колонок, разделенных широкими интервалами. Колончатое распределение нейронов отражает четко фиксированные связи определенных участков ядра и определенных участков улитки. Не отражает ли это способность улитки птиц анализировать частоту?
Изучая развитие слуха и голоса птенцовых птиц в онтогенезе (к этой категории принадлежат и воробьиные, и попугаи), Т. Б. Голубева установила, что слух появляется перед вылуплепием и первоначально характеризуется чувствительностью к звукам низкой и средней частоты до 2 кГц, в последующем происходит расширение воспринимаемого диапазона одновременно с появлением собственной интенсивной вокализации, на последнем этапе происходит дозревание слуховой функции. В соответствии с развитием слуха первыми дифференцируются и развиваются скошенные волосковые клетки, затем низкие и, наконец, высокие. Созревание скошенных клеток завершается к концу интенсивного этапа развития слуха, низких и высоких — на окончательном этапе. Таким образом, по крайней мере у птенцовых птиц развитие слуха в определенном смысле коррелирует с развитием голосообразования, хотя такая корреляция и не является жесткой.
Нет слуховой коры
Наличие слуховой коры отличает слуховую систему млекопитающих от птичьей.
В составе слуховой коры около 1 млн нейронов (слуховой нерв содержит 3·104 нервных клеток, слуховые ядра продолговатого мозга 9·104, ядра среднего и продолговатого мозга по 4·105), определенным образом ориентированных и послойно расположенных. При действии звуковым электродом, погруженным в эту область, удается зарегистрировать биоэлектрические реакции — вызванные потенциалы, отражающие суммарную работу многих нервных клеток.
Большие полушария птиц лишены слуховой коры. Слуховые центры представлены группами беспорядочно расположенных мелких нейронов, не образующих слоев. Этот тип организации центра называется ядерным в отличие от коркового, свойственного слуховой коре млекопитающих (рис. 9).
Слуховые центры больших полушарий птиц открыты совсем недавно. Они были выявлены с помощью методики вызванных потенциалов и специальных гистологических методик. В отличие от млекопитающих таких центров в мозге птиц оказалось несколько, самым первым был открыт центр, расположенный в лобной части полушарий. Все центры располагались не на поверхности мозга, а в его глубине. Они отличались своими характеристиками, и в частности временем возникновения вызванных потенциалов в ответ на действие звука — латентным периодом. Он оказался минимальным у потенциалов лобных центров — 4–6 мс, а в областях, расположенных сзади от них, возрастал до 18 мс.