Рис. 9. Схема проводящих путей и ядер слухового анализатора птиц по результатам электрофизиологических исследований (Harman, Phillips, 1967) 1 — задняя комиссура; 2 — овоидальное ядро; 3 — латеральный лемнисед 4 — дорзальная порция латерального мёзенцефалического ядра
Различие в латентных периодах ответов разных областей полушарий позволяет предполагать, что каждый из них «питается» информацией из разных источников, разных каналов, отличающихся числом переключательных станций. Ведь каждая станция — это задержка ответа полушарного центра по крайней мере на 1 мс.
Поэтому, исследуя латентные периоды различных, предшествующих полушарным центрам уровней, мы можем судить о прохождении слуховой информации от улитки к высшим отделам. Например, латентные периоды ответов слухового нерва кошки составляют 0,8–1 мс, слуховых ядер продолговатого мозга 1,6–1,75 мс, среднего мозга 3–3,5 мс, промежуточного мозга 5–6 мс, слуховой коры 7-10 мс.
Аналогичный ряд мы можем получить и изучая латентные периоды слуховых ядер птиц. Однако здесь нас озадачат короткие латентные периоды лобных центров. Что же, они получают импульсы по какому-то спрямленному пути, в обход промежуточных станций? И каких? На этот вопрос пока невозможно ответить. Во всяком случае, подобного в слуховой системе млекопитающих мы пока не знаем.
Слуховые центры больших полушарий птиц загадочны и непонятны. Первая работа, описывающая вызванные потенциалы лобного центра, была выполнена в Московском университете. Ее авторами были профессор В. И. Гусельников, вьетнамский аспирант До Конг Хунь и один из авторов этой книги. С тех пор прошло 25 лет. Лобные центры исследовались всесторонне и с помощью различных методик. Были высказаны соображения о нескольких восходящих путях, питающих информацией слуховые центры, не только лобный, по и другие. Были изучены основные характеристики центров. И все же мы далеки от решения основных вопросов — как организованы и как работают слуховые области больших полушарии мозга птиц. Хотя бы в первом приближении, в грубом сравнении с млекопитающими. Мы можем предполагать только, что они работают не хуже, раз птицы так хорошо слышат, распознают звуки, используют столь богатый словарь. Но и только.
Характеристики слуха и параметры голоса
Поскольку в звуковом общении участвуют голосообразующне и звуковоспринимающие органы, естественно ожидать определенных корреляций между ними. Следовательно, слух любого вида, а значит, и попугаев, и воробьиных, должен быть настроен на сигналы своих видовых партнеров, отдавая им предпочтение при восприятии. Но что мы видим на самом деле? Обратимся, во-первых, к общему диапазону воспринимаемых частот.
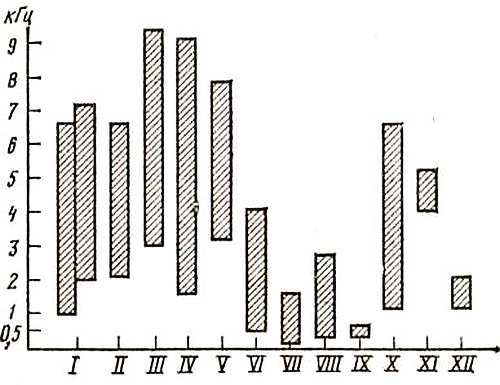
Как показывает рис. 10, верхний порог слуха птиц колеблется у разных видов от 10–12 кГц (голуби, куриные, пластинчатоклювые, дневные хищники) до 16–20 кГц (попугаи, воробьиные, совы, дятлы), однако в сезоны размножения воробьиные птицы способны реагировать на частоты до 35 кГц. Так, американские ученые Фрингс и Кук, используя электрооборонительные методики, установили, что именно до таких частот расширяются верхние границы слуха у скворца в весенние месяцы, тогда как в октябре они снижались до 16 кГц. Пороги слухового восприятия у волнистого попугайчика, определенные в 1940 г. Киехтом, составили 40–16 000 Гц. В этих же экспериментах Киехт определил и другой важный параметр — дифференциальные пороги, которые у волнистого попугайчика оказались близкими к человеческим и составили в области частот 0,3–1 кГц всего 0,3–0,7 % (2–5 Гц), тогда как у кур, например, они оказались равными при частоте 0,3 кГц 3 % (9 Гц), при 1 кГц — 2 % (20 Гц). Голуби же различают только 6 % частоты (в диапазоне 0,3–1 кГц). Значительно более суженной по сравнению с общим диапазоном слухового восприятия является зона повышенной слуховой чувствительности. Как видно из рис. 10, эта зона смещена или в низкочастотную область (голуби, куриные), или занимает область средних частот (воробьиные, попугаи), тогда как у сов она располагается в относительно высоких частотах (до 6 кГц и выше). Именно в области повышенного слуха располагаются самые низкие пороги звукового восприятия, как это показано на рис. 10. Как видно из этого рисунка, кривая слуховой чувствительности волнистого попугайчика несколько отличается от пороговой кривой человеческого слуха, хотя эти различия не слишком значительные.
Рио, 10. Верхний порог слухового восприятия у различных видов птиц (Ильичев, 1972) 1 — пингвин; 2 — кряква; 3 — сокол; 4, 5 — сизый голубь; 6 — серая неясыть; 7 — ушастая сова; 8 — дятел; 9 — волнистый попугайчик; 10 — сорока; 11 — скворец; 12 — клест; 13 — зеленушка; 14, 15 — снегирь; 16 — зяблик; 17 — домовый воробей; 18 — зарянка, а — пороги определялись С помощью поведенческих методик; б — пороги определялись электрофизиологическими методами, белая полоса соответствует зоне оптимального слуха
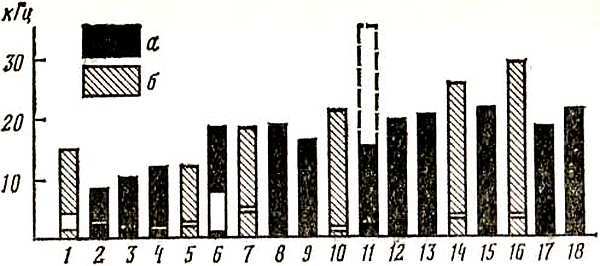
Рис. 11. Спектральный диапазон голоса птиц, имитирующих и не имитирующих человеческую речь 1 — ткачик;· 11 — зонотрихия; 111 — пеночка-теньковка; IV — дрозд; V — певчий воробей; VI — галка; VII — сизый голубь; VIII — курица; IX — тетерев; X — · волнистый попугайчик; XI — домовый сыч; XII — кукушка
Изучая характеристические частоты слуховых нейронов продолговатого мозга волнистого попугайчика, профессор Рурского университета И. Шварцкопф установил, что они стимулируются частотами 0,15-4 кГц, причем большая их часть хорошо реагирует на частоты в области 1,4 кГц.
Если мы теперь обратимся к спектральному диапазону голоса птиц, то обнаружим лишь частичное совпадение его с частотными характеристиками слуха. На рис. 11 изображены общий диапазон голоса и область максимальных частот ряда видов птиц, слуховые возможности которых мы характеризовали выше. Сопоставляя рис. 11 с рис. 10, мы видим, что в большинстве случаев голосовой спектр (нередко и максимальные частоты) укладывается в область повышенного слуха или располагается близко от него. Так, например, крики волнистого попугайчика, охватывающие спектр 1,2–6,4 кГц, вполне укладываются? область его слухового восприятия, голоса сизого голубя и домашней курицы совпадают с областью повышенного слуха. В то же время обращает на себя внимание и значительно избыточная зона слухового восприятия, которую не «занимают» голоса этих видов. Например, слуховой диапазон ушастой совы лишь частично заполнен голосами слетков и взрослых птиц, тогда как большая его часть остается свободной. Как показали исследования одного из авторов этой книги, эта свободная зона, включающая область повышенного слуха, соответствует спектру жизненно важных для совы звуков — писков и шорохов мелких грызунов — полевок и мышей, на которых сова охотится, лоцируя их с помощью слуха.
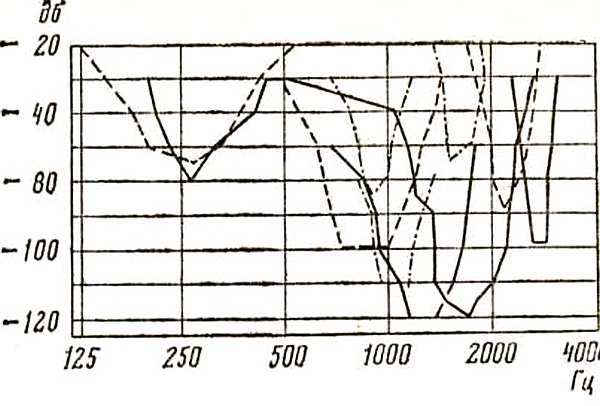
Профессор из ФРГ Иоганн Шварцкопф показал, что рецептивные зоны слуховых нейронов продолговатого мозга располагаются «внутри» диапазона голоса и некоторые из них совпадают с зоной повышенного слуха (рис. 12).
Рис. 12. Настройка слуховых нейронов волнистого попугайчика (Sohwartzkopff, 1957)
Таким образом, отсутствие жесткого соответствия между слухом и собственным голосом обеспечивает резерв, необходимый для восприятия разнообразной и сложной звуковой среды, в первую очередь голосов своих биоценотических партнеров — врагов или жертв, обнаружение и отслеживание которых в пространстве осуществляется с помощью слуха.
Характерно, что и голос человека, который в домашних условиях становится для попугая важнейшим «биоценотическим партнером», обеспечивающим его пищей, водой и этологическим комфортом, также совпадает с зоной повышенного слуха и это совпадение, безусловно, способствует птичьему «говорению».