Детерминация, пожалуй, более четкое понятие. Это такое состояние клеток, когда они уже (часто необратимо) вступили на путь определенной дифференцировки, но внешне сама дифференцировка еще никак не выражается. Молекулярная природа детерминации также неизвестна, но, вероятно, включение определенных генов — если не первый, то непременный этап всякой детерминации. Широкая компетенция эктодермы не позволяет считать ее окончательно детерминированной, хотя при отсутствии особых влияний она дифференцируется в эпидермис. Напротив, компетенция энтодермы так узка, что ее можно называть детерминированной в направлении развития кишечника. Из этого примера ясно, что детерминация — это сужение компетенции. При описании процессов кроветворения чаще употребляют термин «коммитирование». Коммитированной называют клетку, уже осуществившую выбор направления дифференцировки, например в красные или в белые клетки крови.
Между детерминацией и дифференцировкой не может быть четкой границы. Это видно из самого определения детерминации. Оно говорит лишь о том, что доступными нам сейчас средствами мы не видим в клетках проявлений дифференцировки. Может быть, завтра эти проявления мы уже сможем увидеть, например в виде синтеза определенных мРНК. Считать ли их проявлением дифференцировки или условиться называть дифференцировкой только появление особенностей клетки, начиная с синтеза в ней новых белков, — дело вкуса. Поэтому, наверное, правильнее считать детерминацию не процессом, предшествующим дифференцировке, а ее начальным этапом.
И в заключение еще один круг вопросов. В механике развития часто говорили (и иногда используют эти понятия до сих пор) о самодифференцировке и зависимой дифференцировке. В первом случае предполагается, что клетки способны дифференцироваться в каком-либо направлении благодаря их внутренней структуре и свойствам. Как мы увидим дальше, это действительно часто бывает в раннем развитии — целые районы зародыша или отдельные бластомеры исходно различаются особенностями цитоплазмы. На более поздних этапах развития представить самодифференцировку труднее: для того чтобы две действительно одинаковые клетки развивались разным путем, необходимо, чтобы одна из них получила извне какое-то воздействие или информацию, которую не получила другая. Однако и в этом случае, возможно, существует механизм спонтанного вступления на путь дифференцировки, когда отдельные клетки выбирают то или иное направление развития с определенной вероятностью, может быть, под влиянием некоего внутреннего устройства, чем-то напоминающего рулетку.
Однако основным механизмом возникновения различий между клетками являются локальные внешние воздействия. Такая дифференцировка называется зависимой. Эти воздействия могут быть различными — от совсем простых и неспецифических до высокоспецифических, если их вызывают такие соединения, как белки. В настоящее время нам известны десятки примеров зависимой дифференцировки, но, вероятно, ни в одном случае мы не знаем точного механизма таких воздействий. Поэтому мы можем только предполагать, хотя и с большими основаниями, что все локально действующие факторы, вызывающие дифференцировку, в конце концов приводят к активации (включению) определенного набора генов.
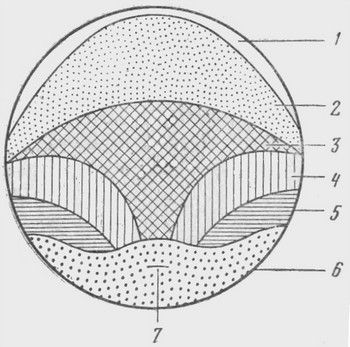
Карта презумптивных зачатков — схема расположения в яйце на стадии ранней гаструлы лягушки материала будущих частей зародыша
Карта могла быть нарисована с большими подробностями, но в действительности на этой стадии между собой различаются только клетки эктодермы, мезодермы и энтодермы. 1 — зачаток эктодермы — будущий эпидермис; 2 — зачаток эктодермы — будущая нервная трубка; 3 — зачаток мезодермы — будущая хорда; 4 — зачаток мезодермы — будущие мышцы спины; 5 — зачаток мезодермы — будущая соединительная ткань, почки, кровь; 6 — зачаток энтодермы — будущий кишечник и его производные (легкие, поджелудочная железа, печень); 7 — бластопор — место, с которого начинается гаструляция: вворачивание энтодермы и мезодермы внутрь зародыша
2. Ооплазматическая сегрегация
Этим термином обозначают возникновение различий между разными частями цитоплазмы яйца (ооплазмы), разделение (сегрегацию) яйца на зоны с несколько различными свойствами. Обычно ооплазматической сегрегацией называют те перемещения компонентов цитоплазмы, которые происходят после оплодотворения и до начала дробления яйца. В действительности же многие процессы ооплазматической сегрегации осуществляются еще в оогенезе, а некоторые продолжаются и в ходе первых делений дробления.
Простейшая ооплазматическая сегрегация происходит в яйцах морского ежа. После четырех делений дробления бластомеры отличаются по способности образовывать некоторые структуры зародыша (султан на анимальном полюсе и кишку на вегетативном). Эта способность меняется вдоль анимально-вегетативной оси. Такое постепенное изменение свойств яйца называют градиентами. У некоторых видов морских ежей эти градиенты проявляются в виде распределения пигментных гранул, располагающихся близко к одному полюсу или образующих кольцо в области экватора.
Более сложно выглядит ооплазматическая сегрегация в яйце амфибий. Еще в ооците создается неравномерное распределение желтка и пигмента вдоль анимально-вегетативной оси: желтка больше на вегетативном полюсе, пигмент покрывает анимальную половину яйца. Ядро оплодотворенного яйца сильно сдвинуто к анимальному полюсу и окружено цитоплазмой, свободной от желтка. После оплодотворения происходит дальнейшая сегрегация: на одной из сторон яйца (она станет спинной стороной) в области экватора возникает более светлая область — серый серп (будущая хордо-мезодерма). Внутри яйца происходят и другие перемещения. В итоге в яйце амфибий можно различать три различно окрашенные зоны: пигментированную анимальную, или эктодермальную; светлую вегетативную, или энтодермальную; экваториальную кольцевую, более широкую в области серого серпа, — мезодермальную. Если до оплодотворения яйцо имело только одну анимально-вегетативную ось, то теперь в нем можно провести и перпендикулярную ей дорсовентральную ось от спины к животу. Две оси образуют плоскость симметрии, которая делит будущий зародыш на правую и левую половины.
В яйце асцидий до оплодотворения никакой сегрегации не обнаруживается: яйцо можно разрезать пополам в любой плоскости и половина, содержащая ядро, образует нормальный зародыш. Такое яйцо изотропно. После оплодотворения в яйце происходят сложные перемещения, которые удается видеть благодаря наличию в цитоплазме различно окрашенных гранул. В итоге в яйце асцидий можно различить не менее четырех зон — анимальную, вегетативную и две серповидные различно окрашенные области, лежащие на уровне экватора друг против друга.
Очень сложные перемещения происходят в яйцах моллюсков. После оплодотворения на вегетативном полюсе образуется так называемая полярная плазма, которая затем движется вдоль поверхности яйца к анимальному полюсу. У некоторых морских моллюсков ооплазматическая сегрегация выражается еще драматичнее: перед первым делением дробления на вегетативном полюсе временно выпячивается особый вырост — полярная лопасть, которая в ходе первого деления достается одному из двух бластомеров, а после второго деления — одному из четырех. В состав этой лопасти попадает полярная плазма и значительная часть митохондрий яйца.
У морских кишечнополостных — гребневиков оплодотворенные яйца имеют у поверхности зеленый слой — эктоплазму, которая в начале дробления равномерно попадает во все бластомеры. Однако в результате последующих делений образуются две группы клеток: одни из них содержат только эктоплазму, другие лишены ее.
Приведенные здесь примеры относятся к тем случаям, когда за ооплазматической сегрегацией можно следить визуально благодаря различной окраске цитоплазматических компонентов. Ho, наверное, чаще — у большинства видов морских ежей, у костистых рыб, птиц и т. д. — ее просто не видно, хотя она, по всей вероятности, существует. Например, яйцо дрозофилы, как и других насекомых, почти не обнаруживает признаков ооплазматической сегрегации, если не считать особых гранул (половых детерминантов), находящихся на заднем конце яйца. Мы уже говорили, как они определяют судьбу ядер, попавших в эту часть яйца. Почти все остальные ядра — после восьми делений их около 250 — перемещаются из центра яйца к его поверхности, где образуют бластодерму. В это время, как было показано экспериментально, определяется их судьба и ядра, попавшие в определенные зоны поверхностной цитоплазмы, образуют клетки так называемых имагинальных дисков. Из каждого диска (они парные) впоследствии образуются определенные органы взрослой мухи (имаго — взрослая). Поэтому можно утверждать, что цитоплазма поверхности яйца неоднородна и что, хотя она вся внешне выглядит одинаково, в действительности в оогенезе происходит ооплазматическая сегрегация, в ходе которой и возникают различные по свойствам зоны цитоплазмы. Некоторые ядра остаются в центре яйца и образуют специальные желточные клетки.