Оказалось, что многие ферменты присутствуют в клетках в нескольких похожих вариантах, их называют изоферментами и различают также по электрофоретической подвижности. Скрещивая два организма, отличающиеся по изоферментам, можно обнаружить, когда в ходе развития впервые появляется отцовский изофермент. Это означает, что включились и проявились отцовские гены данного фермента.
Скрещивание внутри одного вида и использование индивидуальных различий в изоферментах предпочтительны потому, что в этом случае есть уверенность, что оба гена — отцовский и материнский — включаются одновременно. Если же речь идет о межвидовых скрещиваниях или о более отдаленных гибридах, то полной уверенности в этом нет, так как у разных видов стадии, на которых включается тот или иной ген, могут несколько различаться.
Однако некоторые ферменты, принадлежащие к разным видам, электрофоретически не различаются. Многие ферменты трудно или даже невозможно подвергнуть электрофорезу или не удается потом выявить их место в геле с помощью цветной реакции, например такой важный митохондриальный фермент дыхания, как цитохромоксидаза. В составе митохондрий подвергнуть его электрофорезу, естественно, нельзя, а при выделении из них он сразу теряет активность. Для таких случаев в совместной работе нашей лаборатории биохимической эмбриологии (Москва, Институт биологии развития) и лаборатории сравнительной цитологии (Ленинград, Институт цитологии) было предложено использовать различия в теплоустойчивости ферментов.
Действительно, как показали ленинградские ученые В. Я. Александров и Б. П. Ушаков, у хладнокровных животных, обитающих в теплом климате, устойчивость белков к высоким температурам обычно оказывается выше, чем устойчивость аналогичных белков у видов, обитающих в холодном климате. Теплоустойчивость легко измерить, если образцы экстракта белков нагревать при разных температурах в течение определенного времени и потом смотреть процент активности фермента, которая после такого прогрева сохранилась. Температура, при которой сохраняется половина активности фермента, и есть показатель теплоустойчивости.
Первоначально нами совместно с А. А. Кусакиной (Ленинград) было показано, что один из ферментов углеводного обмена — альдолаза у вьюна имеет теплоустойчивость на 5–6° ниже, чем у тропических аквариумных рыб, которых можно скрещивать с вьюном (данио, барбус, разбора и др.). У гибрида вьюна (♀) с данио (♂) (яйца вьюна, сперма данио) на ранних стадиях теплоустойчивость альдолазы была такой же, как у вьюна. Так продолжалось до стадии 23 часа (появление зачатков первых органов), когда теплоустойчивость внезапно стала повышаться. В течение нескольких часов она поднялась на 2,5–3° и далее осталась на том же уровне. Это означало, что именно со стадии 23 часа начинается проявление отцовского гена альдолазы и в результате у гибридов возникает смесь двух ферментов — отцовского и материнского. Когда мы взяли реципрокных гибридных зародышей данио (♀) × вьюн (♂), их теплоустойчивость сначала была высокой, как у данио, а затем на той же стадии 23 часа стала снижаться и также достигла промежуточного значения.
В литературе накопилось довольно много данных о начале экспрессии генов, кодирующих различные ферменты. Такие данные получены в основном методом электрофореза на гибридах лягушек, двух видов форели, курицы и перепела и др. В нашей лаборатории при использовании обоих методов — по электрофоретической подвижности и по теплоустойчивости — на гибридах вьюна и других рыб получены данные о начале экспрессии генов, кодирующих несколько ферментов.
Оказалось, что ни у одного из исследованных нами ферментов гены не начинают проявлять свою активность тогда, когда начинается морфогенетическая функция генов, т. е. на стадии бластулы. Самое раннее проявление отцовских генов было обнаружено для цитохромоксидазы: повышение ее теплоустойчивости у гибридов вьюн × данио было замечено на стадии поздней гаструлы (14 ч развития при 21 °C). Однако некоторые гены, кодирующие такие важные ферменты, как глюкозо-6-фосфатдегидро- геназу или эстеразу, впервые начинают проявляться только в конце эмбрионального развития, а лактатдегидрогеназа еще позже — у ранней личинки.
Во многих случаях, хотя и не во всех, начало экспрессии генов совпадает с началом повышения общей активности фермента. Для эстеразы не только была обнаружена стадия, на которой начинается образование фермента, кодируемого генами зародыша, но и показано, как постепенно после этого исчезает материнский фермент. Однако почему одни ферменты начинают заменяться новыми, кодируемыми генами зародыша относительно рано, а другие — гораздо позже, остается непонятным.
Может быть, это связано со временем жизни самих белков и их мРНК, запасенных в оогенезе. Может быть, на той или иной стадии один изофермент должен заменяться другим. Может быть, быстрый рост отдельных органов зародыша требует соответствующего синтеза ферментов в клетках этого органа. Наконец, рост всего зародыша или личинки предполагает и увеличение количества фермента. Во всех этих случаях может оказаться необходимым появление нового фермента, а для этого необходимо включение собственных генов зародыша.
Подводя итог всей главе, можно сказать, что включение генетического аппарата на определенной стадии развития или включение отдельных генов требует участия специального механизма регуляции. В этом механизме должен быть какой-то датчик стадии развития, запускающий всю цепь событий, в конце которой много генов или один из них начинает транскрибироваться.
В трех последовательных главах мы рассмотрели три «начала» развития — от начала оогенеза, от момента оплодотворения и от начала работы генов зародыша — и убедились, что существенно не то, что называть началом развития, а те последовательно наступающие процессы, которые и составляют раннее развитие. Все они проходят под генетическим контролем. Разница между ними лишь в том, чьи это гены и когда они функционируют. В оогенезе это непосредственный эффект действия материнских генов; при созревании яйца, при оплодотворении и на начальных стадиях развития это отдаленный эффект тех же материнских генов, но опосредованный через запасенные в оогенезе мРНК и белки. Наконец, все последующее развитие происходит снова под непосредственным контролем генов, но уже генов зародыша.
Глава V
Как возникают различия между клетками
Проблема возникновения различий между клетками имеет длительную историю: попытки ее экспериментального решения предпринимались еще в конце прошлого столетия, а теоретические споры преформистов и эпигенетиков восходят к XVII в. Сейчас уже стали известны некоторые важные механизмы, ответственные за дифференцировку клеток зародыша в разных направлениях. Однако до сих пор в этой области больше неясного, а многое еще остается совершенно непонятным.
Этот раздел биологии развития находится почти вне всякой связи с генетикой и молекулярной биологией, а существующие гипотезы носят пока умозрительный характер. Поэтому здесь во многом сохранились чисто феноменологические понятия, т. е. используемые термины являются лишь условными обозначениями того или иного явления. В связи с этим нам кажется правильным начать эту главу с общих представлений, без которых трудно войти в мир экспериментальной эмбриологии, или, как ее еще называли, механики развития.
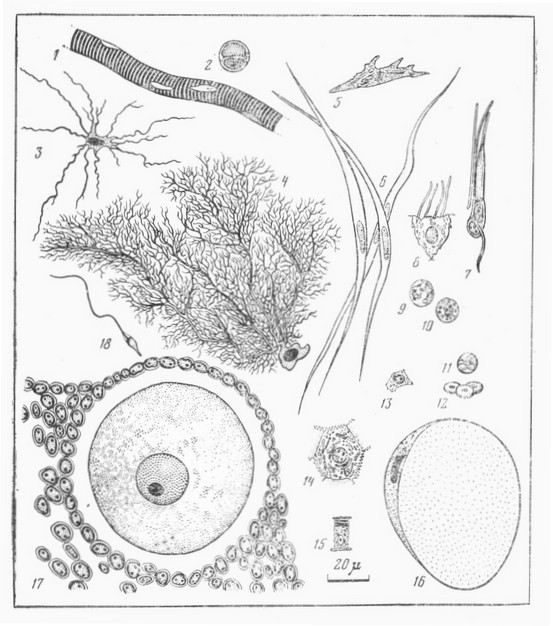
Различия в размерах и морфологии клеток млекопитающих
Все клетки нарисованы при одинаковом увеличении: 1 — многоядерная «клетка» — волокно поперечно-полосатой мышцы; 2 — шванновская клетка, окружающая аксон нейрона; 3 — большая нервная клетка головного мозга; 4 — клетка Пуркинье из мозжечка; 5 — коллагенообразующая клетка соединительной ткани; 6 — клетки гладкой мускулатуры; 7 — палочки и колбочки сетчатки глаза; 8 — клетки реснитчатого эпителия; 9 — полиморфно-ядерные нейтрофилы — один из типов «белой» крови; 10 — антителообразующий лимфоцит; 11 — поперечный срез капилляра, образованного одной клеткой эндотелия, в просвете видны эритроциты; 12 — эритроциты; 13 — клетка хряща; 14 — клетка печени; 15 — слизистая клетка желудка; 16 — жировая клетка соединительной ткани, почти целиком заполненная жиром; 17 — ооцит и окружающие его фолликулярные клетки; 18 — сперматозоид