Обсуждение этого вопроса завело бы нас слишком далеко. Поэтому вернемся к синтезу менделевской генетики и классического дарвинизма. Четвериков в 1926 году показал, что именно те ограничения, которые отличают харди-вейнберговскую популяцию от реальной, и делают возможным процесс эволюции. Рассмотрим же факторы эволюции по Четверикову.
Как возникает разнообразие
Из предыдущих глав вы уже знаете, что гены не абсолютно стабильны. Меняется по целому ряду причин структура отдельных генов, их местоположение в хромосомах, и сам хромосомный набор претерпевает всевозможные изменения — вплоть до многократного умножения. Судьба этих изменений оказывается различной.
Многие из них приводят к появлению нежизнеспособных форм, погибающих на той или иной стадии развития. Другие как бы воздвигают вокруг носителей таких мутаций «китайскую стену» нескрещиваемости. В большинстве случаев подобные отщепенцы сходят с жизненной арены, оставляя бесплодное потомство. Наконец, есть и такие, которые включаются в генный фонд и передаются из поколения в поколение. Они претерпевают при каждом мейозе все перипетии генетической рекомбинации — и в конечном счете рано или поздно, оказываясь в гомозиготном состоянии, проявляются в фенотипе. Вид на протяжении своей истории непрерывно обогащается мутантными аллелями. По образному выражению Четверикова, вид, как губка, всасывает мутации (он, правда, предпочитал другой термин — геновариации, в практике не прижившийся).
Чем же, однако, объяснить тот факт, что виды на протяжении многих поколений остаются стабильными, сохраняют, как говорят, «дикий тип»? Напомним, что большинство мутаций рецессивно. Каждая мутация — явление достаточно редкое; лишь после того, как она размножится в последующих поколениях потомков, вероятность встречи двух одинаковых генов в зиготе возрастет. Селекционер может ускорить этот процесс: практически все породы домашних животных и сорта культурных растений гомозиготны по разным рецессивным генам. Скрестив два разных сорта их, мы нередко получаем в потомстве возврат к прежнему «дикому типу». Это знал и Дарвин, скрещивавший голубей разных пород и получавший в потомстве «сизарей». Механизм здесь тот же, что и в случае супрессорных мутаций, о которых мы уже упоминали, — дефект одного генома исправляется другим.
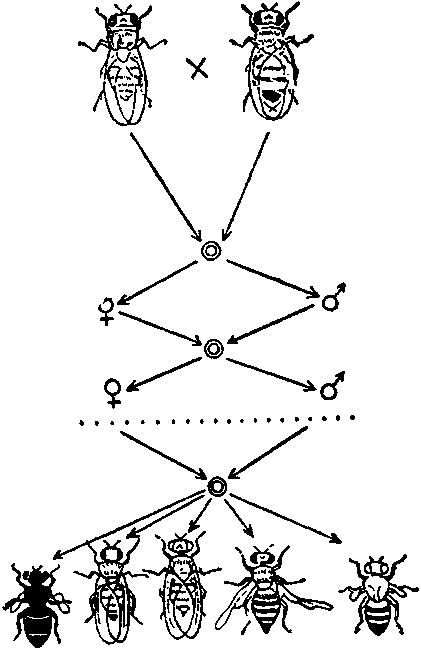
Схема опыта С. С. Четверикова. В потомстве одной дрозофилы после ряда поколений близкородственного разведения выщепляются рецессивные мутантные гомозиготы, до того в скрытой виде находящиеся в однообразном, «диком типе».
При внешнем фенотипическом однообразии геномы видов настолько обогащены скрытым разнообразием, что, остановись мутационный процесс, стань гены абсолютно стабильными, эволюция все равно не прекратится. Реализовать в фенотипах уже накопленное генетическое разнообразие в сколько-нибудь обозримые сроки невозможно.
Из построений Четверикова вытекает важное следствие. Наибольшее значение для эволюции имеют лишь те изменения генетического материала, которые, находясь в рецессиве, не проявляются в фенотипе. Не подвергаясь действию отбора, они могут накопиться в популяциях в достаточном количестве. Из этого правила есть, разумеется, исключения. Бактерии и сине-зеленые водоросли, то есть организмы с неоформленным ядром, не имеющие фазы мейоза, гаплоидны. Они имеют только один набор генов, поэтому каждая мутация немедленно проявляется в фенотипе, у них нет запасов генетической изменчивости. Образно выражаясь, они не помнят прошлого и не готовятся к будущему, живя сегодняшним днем. Бурный темп их размножения позволяет им выстоять в жизненной борьбе. Однако они, приспособляясь, не эволюционируют, оставаясь и в наши дни практически теми же, как и в то время, когда были единственными обитателями нашей планеты. Недаром и бактерии, и даже вирусы выработали свои, оригинальные механизмы генетической рекомбинации, которым, однако, далеко до мейоза и полового процесса высших организмов.
Не каждая мутация, вновь возникнув, может закрепиться в потомстве. Ведь не каждый ее носитель доживает до половозрелости. Существует, помимо накопления мутаций, обратный процесс их потери, обеднения генного фонда. Вероятность потери мутации можно рассчитать: если мы примем, что из потомства одной самки выживают двое, то есть численность популяции не увеличивается и не уменьшается (то, что немцы называют система двух детей — Zweikinder-system), то примерно 37 шансов из ста за то, что эта мутация будет потеряна[9]. В создании генного фонда популяции участвуют две противоборствующие тенденции.
Можно ли утверждать, что мутационный процесс наряду с генетической рекомбинацией сможет обеспечить нормальный ход эволюции, то есть возникновение новых видов, приспособленных к окружающей среде? Напомним, что аналогичные идеи развивал Де Фриз. Однако считать возникновение генетического разнообразия единственным двигателем эволюции нельзя. Мутационный процесс равнодушен к судьбам организмов, геномы которых он изменяет. Приспособленность к условиям внешней среды он создать не может.
«Волны жизни», «горлышко бутылки» и «принцип основателя»
Наряду с мутационным процессом второй фактор эволюции по Четверикову — колебания численности популяций, которые он образно называл «волнами жизни».
Каждый из нас наблюдал «волны жизни» воочию. Ранней весной редко можно увидеть случайную перезимовавшую между оконными рамами муху, а в августе мы страдаем от их назойливости. Год на год не приходится — в одно лето нет житья от комаров, а в другое они редки. Вспышки численности некоторых видов имеют воистину планетарный характер. Таковы, например, массовые миграции саранчи в годы ее усиленного размножения, годы «мышиной напасти», путешествия многочисленных стад полярных пеструшек — леммингов.
Некоторые из этих вспышек цикличны: они имеют сезонный, годовой или многолетний период (часто, хотя порой и без достаточного обоснования связываемый с солнечной активностью). Другие же апериодичны. Это вспышки численности организмов, попавших в новую среду обитания без достаточного количества врагов, эпидемии гриппа и иных заболеваний, с быстротой пожара распространяющиеся по планете нашествия вредителей из других стран.
Колорадский жук у себя на родине, в Северной Америке, никогда не достигал высокой численности, питаясь дикими растениями из семейства пасленовых. Печальную славу злостного вредителя он завоевал на европейских картофельных полях. То же можно сказать и об американской виноградной тле — филлоксере. Континент Евразии не остался, впрочем, перед Америкой в долгу, «наградив» ее непарным шелкопрядом, воробьями и длинным списком сорняков. Вспомним также нашествие кроликов и кактусов опунций на Австралию.
Отнюдь не всегда такие подъемы численности видов вредоносны. Бурное развитие в Каспийском море акклиматизированного там азовского червя — нереиса значительно увеличило кормовую базу рыб. Это в свою очередь компенсировало оскудение Каспия, наступившее в результате зарегулирования стока Волги. Не будь нереиса, осолонение Северного Каспия нанесло бы каспийским рыбам непоправимый ущерб. Никто не может возразить и против «нашествия» на нашу страну американской мускусной крысы — ондатры, расселившейся от Камчатки до Мурмана, от северной Сибири до Средней Азии (правда, по некоторым данным, она вытесняет ценного зверя — выхухоль). И все же испортить неразумным вселением сложившуюся структуру сообществ растений и животных гораздо легче, чем улучшить ее. Нелишне напомнить, что акклиматизации нереиса в Каспии предшествовала многолетняя работа. Сейчас с развитием авиации, облегчающей перевозки, порой случается и такое, что сначала акклиматизируют, а потом скромно уступают друг другу честь этого замысла. Дело доходит до курьезов, когда вместе с ценными объектами (а то и вместо них) завозят совсем нежелательные. Так, вместе с ценными растительноядными рыбами в Среднюю Азию завезли головастиков чернопятнистой лягушки с Дальнего Востока (уж этого-то можно было избежать!).