Все эти линии на самом деле вымерли, что, конечно, не противоречит предположению об инадаптивном характере данных тенденций. Правда, вымирание филетических линий — дело обычное. Ведь вымерли же, например, все виды семейства Hominidae, за исключением Н. sapiens, и вообще филетический пучок высших приматов не выглядит преуспевающим. Не станем же мы говорить об инадаптивном сверхразрастании головного мозга у гоминид. Кроме того, отметим, что все отмеченные выше случаи «сверхразвития» рабочих структур противоречат принципу симморфоза, согласно которому ни один орган не бывает развит свыше потребности в нем в моменты максимальных нагрузок.
Однако вернемся к правилу Копа. Проследим в какой-нибудь филетической группе за динамикой видовых распределений по величине особей. На рис. 17, а приведены распределения по диаметру раковины у брахиопод отряда Rynchonellida на трех этапах их исторического развития. Аналогичную картину демонстрируют и аммонитины юры (рис. 17, б). Первое, что бросается в глаза, — это увеличение размаха распределений, обусловленное прежде всего появлением видов с большими размерами особей. Кроме того, эти распределения асимметричны — с длинным «хвостом» в правой части. Очевидно, что среднее значение (центр тяжести) распределений также сдвигается с ходом времени вправо. Отметим, что виды с мелкими особями продолжали существовать всегда, хотя их доля постепенно убывала.
Сходную картину можно наблюдать и для видовых распределений по признакам, величина которых хорошо коррелирует с массой тела. Чтобы понять это макроэволюционное явление, сначала рассмотрим, как вообще формируются распределения видов по количественным признакам. Отметим, что после перехода к логарифмической шкале измерения признака многие из этих распределений симметризуются, становясь близкими к нормальным.
Межвидовые дистанции
Мы на глаз, без труда различаем млекопитающих, принадлежащих разным родам. Например, волка и лисицу, рысь и леопарда, козу и овцу и т. д. Еще легче разобраться с представителями разных семейств, отрядов и других таксонов высокого ранга. Сортировка видов одного рода требует большего внимания, хотя в случае млекопитающих она обычно удается, вероятно, помогает собственная принадлежность к данному классу. Видимо, поэтому мы можем отличить друг от друга льва, тигра и леопарда, несмотря на то, что все они входят в один род Panthera. Кстати, отличить череп льва от черепа тигра под силу только специалисту, а гибрид этих самых сильных кошек — тигролев — вполне жизнеспособен. Куда труднее различить родственные виды у насекомых, особенно когда речь идет о так называемых видах-двойниках.
Человек при оценке степени морфологического сходства пользуется присущей ему интуитивной способностью распознавать образы. «Ненаучность» такого метода очевидна. Первая вполне научная система организмов была создана К. Линнеем в 1735 г. Она основана на сопоставлении рядов четко регистрируемых морфологических признаков, которыми можно охарактеризовать типичную особь. Чем больше признаков совпадает в таких рядах, тем выше степень сходства сравниваемых видов. Чем меньше число совпадений, тем выше ранг таксономической дистанции между видами. Слабость этого подхода заключается в субъективном характере выбора признаков, положенных в основу системы.
К сожалению, более объективные молекулярные методы обычно дают информацию только о времени эволюционной дивергенции видов. Дело в том, что удобные для регистрации различия в строении белков или нуклеиновых кислот чаще всего обусловлены нейтральными аллельными заменами, которые фиксируются с более или менее постоянной скоростью — скоростью мутирования (см. (3.26)).
В настоящее время существует ряд способов измерения так называемой морфологической (фенотипической) дистанции между двумя группами особей. Несмотря на различия в математическом оформлении, все эти оценки являются сложным отражением того, во сколько раз (или на сколько процентов) отличаются значения одних и тех же признаков у сравниваемых групп особей. По-видимому, из-за этого оцененные разными способами значения морфологических дистанций неплохо коррелируют друг с другом.
Многочисленные измерения фенотипических дистанций позволяют прийти к некоторым выводам, довольно существенным для нашей темы. Во-первых, для многих совсем не родственных видов животных средние расстояния между популяциями одного вида обычно весьма близки, и, во-вторых, для морфологических дистанций, отделяющих виды одного рода, также характерен свой средний уровень, который, как правило, в 1,5–2 раза превосходит межпопуляционный. Эти результаты наводят на мысль, что превращение популяций в виды сопряжено с достижением их особями вполне определенного значения относительного сдвига величины ряда количественных признаков. Что же происходит с видом в момент видообразования? Очевидно, приспособление к новым экологическим условиям.
Хорошо известно, что для каждого вида ряд жизненно важных параметров среды не должен выходить за пределы некоторых критических значений. Так, для жизни рифообразующих коралловых полипов необходимо, чтобы температура воды не опускалась ниже 20,5 °C. Полный комплекс требований вида к параметрам среды может быть объединен понятием экологической ниши вида. Таким образом, каждому виду соответствует его экологическая ниша. В одной нише устойчиво может существовать только один вид (принцип Гаузе), если вдруг по какой-то причине в ней окажутся два, то со временем один из них уступит место другому, скорее всего, он вымрет. Филетическое выживание вида (видообразование) сопряжено с его переходом в соседнюю экологическую нишу, при этом же, очевидно, происходят сдвиги значений многих количественных признаков особей. Мы уже отмечали (см. параграф «Аллометрия» в гл.2), что сдвиги подобного рода обычно связаны с изменением абсолютных размеров структур и прежде всего с изменением средней массы животного.
Знаменитый американский эколог Дж. Хатчинсон предположил, что для завершения экологической дифференциации средняя величина морфологического признака, непосредственно ответственного за адаптацию, должна измениться в какое-то фиксированное число раз. Оказалось, что довольно часто выполняется одно удивительное правило: массы особей родственных видов животных, обитающих в одной местности (т. е. в сходных экологических условиях), отличаются друг от друга в число раз, примерно кратное двум. Как будто для экологического разобщения особи таких видов должны различаться по массе примерно в 2 раза. Из этого правила Хатчинсона следует, что линейные размеры тела отличаются минимум в 21/3 ≈ 1,26 раза, или, грубо, на 30 %. Особенно четко это правило выполняется для размеров пищедобывающих структур животных. Прямые измерения показывают, что чаще всего линейные размеры этих структур (L) отличаются, как минимум, на 28 %. Отсюда вытекает, что размер L можно представить как некую минимальную величину Lmin, умноженную на произведение ряда сомножителей хi, близких к 1,28, т. е.
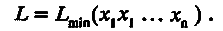
(5.1)
После логарифмирования этого выражения имеем
(5.2)
Заметим, что ∑lnxi представляет собой сумму близких по порядку независимо варьирующих величин, поэтому распределения большого числа видов по логарифму размеров или массы особей должны быть близкими к нормальным.
Формирование видовых распределений
Построим простейшую модель видовых распределений. Будем считать, что для экологической дифференциации видов их особи должны отличаться по размерам на 28 %, соответственно натуральный логарифм их размеров — приблизительно на 0,25. Разобьем горизонтальную ось, вдоль которой изменяется логарифм линейных размеров, на интервалы шириной 0,25. Пусть вид-основатель филетической группы по размеру своих особей находился в интервале х0. Через единичный отрезок времени (через один шаг модели) он произведет два дочерних вида, которые попадут в соседние интервалы х-1, и х1. Отметим, что интервал х0 остается занятым материнским видом. В следующий временной отрезок каждый из трех видов приступит к «завоеванию» своих соседних интервалов. Продолжая такой процесс, мы вскоре получим довольно гладкое симметричное (биномиальное) распределение, очень близкое к нормальному. С каждым шагом оно расширяется, его дисперсия увеличивается пропорционально возрасту таксона, а сигма следует за квадратным корнем из этого возраста. Однако центр тяжести распределения (его среднее значение) должен оставаться на месте, точно соответствуя величине признака у вида-основателя филетической группы — х0 (рис. 18, а).