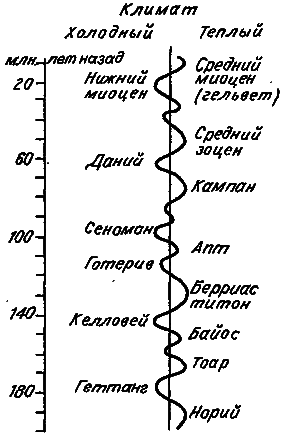
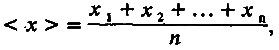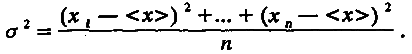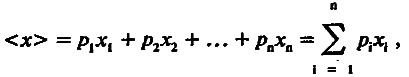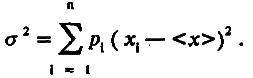Итак, данные геологии свидетельствуют о том, что на Земле действительно происходили и происходят очень медленные и однонаправленные изменения среды, захватывающие огромные территории. Характерное время таких изменений выражается миллионами лет.
Рис. 12. Изменение среднегодовой температуры в течение юры и мела (по: [Красилов, 1977]).
Легко представить, что чем меньше ареал, занимаемый видом, тем больше у него шансов попасть под действие рассматриваемых здесь медленных однонаправленных сдвигов среды. Американский палеонтолог А. Буко проследил за изменением в течение силура и девона видового состава брахиопод, обитавших в разных геологических провинциях, по-видимому, просто в разных эпиконтинентальных морях. Оказалось, что единственным фактором, от которого зависела скорость изменения фауны, была площадь этих морей. Чем меньше площадь, тем быстрее изменялся состав брахиопод. Заметим, что речь идет о миллиардах особей, занимавших ареалы в миллионы квадратных километров.
Рассмотрев в этой главе обстоятельства, при которых возможны ненейтральные изменения генофонда вида, мы приходим к двум важным выводам: во-первых, уровень приспособленности вида к среде следует признать всего лишь удовлетворительным, и, во-вторых, скорость фенотипического сдвига (приспособительного характера) не может быть очень высокой.
Глава 4. Ответ на отбор
Лорд Сомервиль, упоминая о том, чего животноводы достигли по отношению к овце, говорит: «Кажется, будто они начертили на стене форму, совершенную во всех отношениях, и затем придали ей жизнь».
Ч. Дарвин
Обычно после прочтения книги Ч. Дарвина «Происхождение видов…» и даже после ознакомления с трудами крупнейших неодарвинистов от А. Вейсмана до Э. Майра у читателя остается впечатление, что отбор может все. Ограничения на его творческие возможности накладывают лишь разного рода экологические перипетии, но будь обстоятельства благоприятны, отбор за считанные миллионы лет мог бы в буквальном смысле создать из мухи слона.
Со времен Дарвина в качестве главного аргумента в пользу безграничных возможностей отбора выдвигаются успехи человека в создании хозяйственно ценных форм растений и животных. Особенно впечатляют результаты методической селекции последних двух столетий. Чаще всего такая селекция была направлена на повышение мощности вполне конкретной рабочей структуры. Чем не модель для изучения прогрессивной эволюции? Заметим, что скрещивая более и менее продвинутые на пути такого «прогресса» формы и следя за их потомством, можно было бы вскрыть генетическую подоплеку мощности рабочих структур. Итак, если мы действительно хотим проверить тезис о всесильности отбора и научиться измерять его творческие возможности, нам следует совершить небольшой экскурс в генетику количественных признаков.
Изменчивость особей
Вооружившись подходящим измерительным инструментом, мы можем убедиться, что взрослые особи любой природной популяции отличаются друг от друга по множеству характеристик (признаков): размерам тела и органов, частоте пульса, скорости бега, численности потомства и т. д. Попробуем найти способ, как оценить степень изменчивости популяции по таким мерным признакам.
Выберем наугад большое число особей (желательно одного возраста и пола) и измерим у них какую-нибудь характеристику. Ясно, что в любой выборке можно найти одну особь с наименьшей величиной признака и одну — с наибольшей. Разность между этими значениями — диапазон варьирования мерного признака — может служить грубой мерой его изменчивости в популяции. Разобьем диапазон варьирования на ряд равных по величине интервалов. Очевидно, что с помощью этой процедуры мы разделим свою выборку на классы, каждый из которых формируется особями со значением признака, не выходящим за пределы соответствующего интервала. Теперь всю эту совокупность особей можно охарактеризовать ее распределением по признаку, т. е. числом особей, входящих в каждый класс. Фактически мы задаем тем самым математическую функцию численности особей от величины измеряемого параметра.
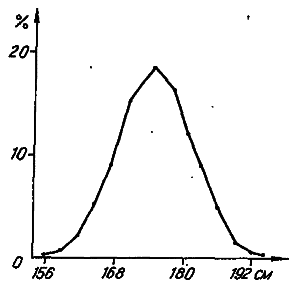
Рис. 13. Распределение студентов Гарвардского университета по росту (no: [Castle, 1916]).
Эту функцию удобно воспринимать графически (рис. 13). На оси абсцисс откладываем классовые интервалы, а ордината отражает численность особей, «приписанных» к соответствующему интервалу.
К настоящему времени изучены распределения по всевозможным признакам у большого числа популяций разных видов. Оказалось, что их графики обычно представляют собой непрерывные (гладкие) кривые с одной вершиной. Признаки, имеющие такие гладкие одновершинные распределения, в генетике принято называть количественными. Нередко графики этих распределений имеют симметричную колоколообразную форму, напоминая в этом отношении нормальное, или гауссовское, распределение, давно известное в теории вероятностей.
Центральным объектом теории вероятностей является так называемая случайная величина, которая характеризуется своим распределением, т. е. вероятностью принимать значения в заданных интервалах. В этой области математики доказывается, что нормальное распределение возникает, когда случайная величина является суммой большого числа независимо варьирующих, но близких по величине слагаемых. Классический пример представляет собой распределение ошибок, изученное великим немецким математиком К. Гауссом.
Вся информация о конкретном нормальном распределении заключена в двух его параметрах — среднем значении и дисперсии. Их можно легко вычислить и для нашей выборки особей. Среднее значение <x> — это просто среднеарифметическая величина признака. Угловые скобки обозначают операцию усреднения. Итак,
(4.1)
где х1, х2…, хn — величина признака у первой, второй и т. д. особей; n — число особей в выборке.
Дисперсия распределения σ2 отражает изменчивость особей по величине признака и является, по определению, средним квадратом отклонения величины признака от его среднего значения:
(4.2)
При получении распределения особей по признаку мы разбиваем всю выборку на n классов, каждый из которых может быть охарактеризован численностью и средним значением признака. Для удобства сравнения разных распределений вместо численности класса лучше взять долю, которую составляют его особи в выборке. Таким образом, i-й класс можно охарактеризовать его долей (рi) в выборке и средним значением признака (хi). В этом случае среднее значение и дисперсия вычисляются по формулам
(4.3)
(4.4)
Компоненты дисперсии
Теперь попробуем разобраться в причинах изменчивости особей в природных популяциях. Практический опыт подсказывает, что это явление зависит от среды и от наследственности. То, что среда (в частности, качество питания) влияет на величину количественных признаков, знают все. Влияние наследственности также хорошо известно, достаточно напомнить поговорку: «Яблоко от яблони недалеко падает».
Действие на признак обоих факторов (среды и наследственности), по существу, независимо, что весьма упрощает проблему разложения наблюдаемой (фенотипической) изменчивости на компоненты, связанные с действием каждого фактора. В теории вероятностей доказывается, что дисперсия суммы независимо варьирующих случайных величин является суммой дисперсий каждой из них. Соответственно и фенотипическую дисперсию по величине любого признака (σ2) можно разложить на два слагаемых, на две компоненты — средовую (σе2) и генотипическую (σg2) дисперсии: