Предположим, что вероятность вымирания определяется одним внутренним свойством линии — генетически определяемой способностью преодолевать долговременные, неблагоприятные для нее изменения среды. Назовем это свойство адаптируемостью. Как закодированное в геноме оно должно наследоваться при видообразовании, т. е. передаваться (с легкими изменениями) от вида-предка к виду-потомку. Следовательно, виды-соседи по филетической цепи и даже виды из разных линий одного филетического пучка должны обладать близкими значениями адаптируемости.
Виды с высоким значением адаптируемости имеют больше шансов преодолеть неблагоприятные изменения среды, поэтому средняя адаптируемость у видов-потомков должна быть несколько выше, чем у видов-предков. За время существования многоклеточных (700–800 миллионов лет) произошло около ста довольно крупных изменений фауны, которые могут служить границами ярусов — стратиграфических аналогов геологических веков. Можно предположить, что филетические линии, выдержавшие в конце концов все эти испытания, отличаются от вымерших повышенной скоростью преобразования наследственной информации.
На филетический пучок можно взглянуть как на популяцию бесполых организмов. В такой популяции каждая особь (отдельный вид) через фиксированный промежуток времени (равный продолжительности жизни вида) или гибнет, или оставляет потомков (дочерние виды). При этом адаптируемость вида — это приспособленность отдельной особи в такой популяции, т. е. адаптируемость можно определить как вклад данного вида в филетический пучок на следующем отрезке макроэволюционного времени. Заметим, что длительность такого отрезка соизмерима с миллионом лет.
Как показал в 1955 г. американский генетик Дж. Кроу, в бесполой популяции идет постоянное возрастание средней приспособленности. При этом скорость данного процесса в точности равна популяционной дисперсии по приспособленности. Поскольку дисперсия (как квадрат сигмы) не может быть отрицательной величиной, то средняя адаптируемость филетического пучка в ходе эволюции может только возрастать. Не является ли это свидетельством эволюционного прогресса?
Эволюционная адаптация к новой среде предполагает фиксацию серии благоприятных мутаций. В соответствии с уравнением (3.31) скорость этого процесса прямо зависит от размера популяции, скорости появления благоприятных мутаций и степени их благоприятности. Мы определили адаптируемость как внутреннее, т. е. заключенное в геномах особей, свойство вида. Из перечисленных выше факторов лучше всего ей соответствует скорость появления благоприятных мутаций. Но процесс мутирования слеп, поэтому из всех возникающих мутаций лишь небольшая их часть может быть благоприятной в данной конкретной (неблагоприятной для популяции) обстановке. Следовательно, для повышения адаптируемости было бы неплохо увеличить скорость появления мутаций, способных компенсировать изменения среды, наиболее частые в экологическом пространстве вида.
Поскольку мы ввели адаптируемость как неотъемлемое свойство генома типичной особи вида, то тем самым признали, что могут быть мутации как уменьшающие, так и увеличивающие адаптируемость. Фактически мы предположили, что могут существовать мутации, изменяющие скорость появления благоприятных мутаций. На первый взгляд кажется, что для этого достаточно просто повысить мутабильность среднего гена, однако такое решение вопроса весьма сомнительно. Ведь рост мутабильности автоматически означает увеличение генетического груза, т. е. падение уровня приспособленности популяции. Наиболее перспективно выглядят мутации, повышающие долю благоприятных среди всех мутаций, возникающих в геноме за поколение. Если такие мутации, какими бы фантастичными они ни казались, действительно существуют, то они должны накапливаться в генофондах видов при браковке филетических линий по адаптируемости.
В настоящее время существуют две точки зрения на филетическое видообразование. Одна традиционная, градуалистская, впервые высказанная Дарвином, предполагает, что нет четкой границы, где один вид сменяет другой, поэтому сдвиг фенотипа средней особи идет постоянно, хотя в его скорости возможны значительные колебания. Согласно другой, высказанной С. Гоулдом и Н. Элдриджем в 1972 г., вид в течение миллионов лет не изменяется, находясь в состоянии морфологического «стазиса». Затем за относительно короткий промежуток времени происходит его превращение в новый вид — видовая трансформация. Хотя этот «краткий» миг может длиться и несколько десятков тысяч лет, все равно его длительность ничтожна в сравнении с миллионами лет стазиса.
Если верна градуалистская модель, то процесс адаптации к новым условиям длится миллионы лет, и все это время сохраняются условия для фиксации мутаций, увеличивающих адаптируемость. Пунктуалистская модель предполагает, что отбор в пользу таких мутаций действует наверняка только в период видовой трансформации, и ценные для этого момента аллели могут быть утрачены во время стазиса, если они не вносят вклад в относительную приспособленность особей. Следовательно, аллели, благоприятные для филетического выживания вида, должны быть благоприятными и в микроэволюционном смысле, т. е. повышать уровень относительной приспособленности их носителей.
Заметим, что, введя представление об адаптируемости, мы получили, как следствие, макроэволюционную тенденцию к ее росту. Эту тенденцию можно было бы интерпретировать как накопление эволюционирующей филетической группой информации о неблагоприятных факторах, действующих в ее экологическом пространстве (адаптивной зоне). Как подойти к анализу подобного рода информации, вероятно, запечатленной геномом? Скорее всего такая генетическая запись должна каким-то образом отразиться на фенотипе особей. Поэтому и тенденция к росту адаптируемости в ходе эволюции филетической группы могла бы выглядеть как долговременная тенденция к изменению в определенном направлении морфологии особей ее видов. В связи с этим встает вопрос, не являются ли хорошо известные палеонтологам долговременные тенденции отражением процесса возрастания адаптируемости.
Макроэволюционные тенденции
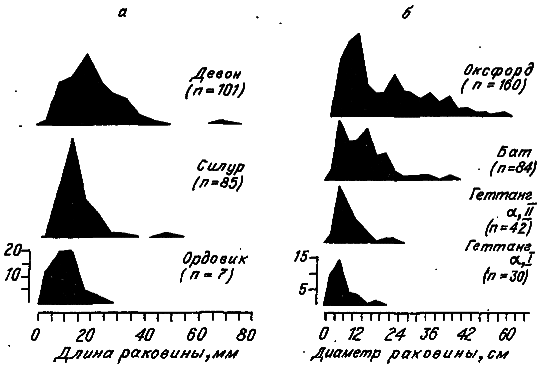
Крупнейший вклад палеонтологии в наши знания об эволюции живых организмов состоит в обнаружении во многих филетических группах особых очень долговременных тенденций к сдвигу в одном направлении некоторых количественных признаков. Пожалуй, самой известной из них является тенденция к увеличению размеров индивидов — так называемое правило Копа. Значительный эмпирический материал свидетельствует, что обычно зоологические таксоны крупных животных происходят от видов с относительно небольшими особями. Между прочим, род Homo происходит от гораздо более мелких обезьян.
Э. Коп объяснил свое правило как классический ламаркист (кстати, основоположник психоламаркизма). Он считал, что в основе открытой им тенденции лежит пресловутая «ростовая сила», стремящаяся под действием психики животного увеличить его размеры. Такой филогенетический рост тела протекает беспрепятственно в периоды геологического затишья и сопровождается разрастанием многих морфологических структур. Тенденции противостоит процесс вымирания гигантов и монстров при резких сменах климата.
Рис. 17. Изменение видовых распределений брахиопод (а) и аммонитин (б) по размеру раковины в историческом развитии (по: [Stanley, 1973]).
Отметим одну особеннность коповского ламаркизма: внутренние стремления индивида снижают адаптируемость филетической линии и влекут последнюю к гибели. Любопытно, что включение в эволюционный сценарий особых роковых факторов — довольно обычное дело для теоретиков-эволюционистов. Достаточно вспомнить о трех таких драматических историях: 1) самоубийственной тенденции юрских устриц (грифей) к созданию нераскрывающихся раковинных створок; 2) инадаптивном сверхразрастании непомерно тяжелых рогов у ирландского оленя; 3) развитии сверхмощных клыков, затрудняющих кусание, у саблезубых кошек.