А = S*σ*Δl, (2.6)
где S — площадь поперечного сечения мышцы; σ — сила, развиваемая мышечным пучком единичного сечения.
Обширный экспериментальный материал свидетельствует, что у всех животных величина σ очень близка. Это объясняется универсальностью строения элементарного мышечного волокна (миофибриллы) и тождественностью молекулярного механизма его сокращения. Поскольку гомологичные мышцы в среднем укорачиваются на одну и ту же долю (примерно на 30 %), то чем мышца длиннее, тем на большую длину она сокращается. Следовательно, работа, совершаемая мышцей при сокращении, пропорциональна ее сечению и длине, т. е. пропорциональна ее объему (или массе).
Теперь посмотрим, как все это отражается на двигательной активности животного и прежде всего на его беге. Бег состоит из последовательности циклов сокращения и расслабления практически всех скелетных мышц. При каждом таком цикле животное перемещается на один шаг, совершая работу, пропорциональную массе своей мускулатуры. А так как эта последняя у млекопитающих составляет в среднем около 40 % от массы тела, то получается, что работа по перемещению на один шаг пропорциональна массе животного, т. е.
Аш~ М. (2.7)
Очевидно, что чем длиннее ноги у животного, тем больше его шаги, т. е. тем меньше нужно их сделать для преодоления одной и той же дистанции. Как показывают измерения, у млекопитающих разных видов длина ног пропорциональна M0,33. Значит, число шагов (n), нужное для покрытия фиксированного расстояния, скажем, один километр, должно быть связано с массой тела соотношением
n ~ М-0,33. (2.8)
При этом животное совершает работу A, равную nAш, т. е., учитывая (2.7) и (2.8),
A ~ М0,67. (2.9)
Поделив обе половины этого соотношения на М, получим, что работа по транспорту единицы массы тела на фиксированное расстояние (A0) должна уменьшаться при укрупнении животного, точно отражая сокращение необходимого числа шагов:
А0 ~ М-0,33. (2.10)
Объем работы А0 можно оценить по количеству кислорода, необходимому для переноса килограмма массы тела на один километр. Оказалось, что эта величина ц — кислородная цена транспорта единицы массы — падает с увеличением размеров животного, подчиняясь аллометрическому уравнению
ц = c*M-0,33. (2.11)
Ясно, что кислородная цена транспорта всей массы животного на один километр (Ц) будет в М раз больше, т. е.
Ц = с*М0,67. (2.12)
Самая поразительная особенность этого уравнения заключается в том, что оно справедливо для всех бегающих наземных животных — ящериц, млекопитающих и даже насекомых. Значение коэффициента с изменяется у разных групп очень незначительно — от 0,6 до 0,8 литра кислорода для перемещения одного килограмма на один километр. Фактически это означает, что эффективность локомоторного аппарата у всех наземных животных примерно одна и та же, вне зависимости от их эволюционной продвинутости. Скорее всего это постоянство зиждется на единстве строения миофибриллы — основной рабочей структуры любой скелетной мышцы.
Теперь посмотрим, как связана с массой тела мощность, развиваемая животным при беге. Напомним, что мощность — это работа, совершаемая за единицу времени. Заменив в уравнении (2.6) величину Δl на абсолютную скорость укорочения мышцы v, мы получим формулу для оценки мощности мышечного сокращения (Pm):
Pm = S*σ*v. (2.13)
Поскольку максимальные значения v для мышц разных видов животных (при одной температуре) близки, то максимальная мощность мышцы определяется площадью ее поперечного сечения S. Эта площадь при изометричном росте должна увеличиваться пропорционально М0,67, и точно так же должна возрастать мощность мышечного сокращения. Но, как мы знаем (см. уравнение (2.3)), мощность метаболизма увеличивается с ростом массы существенно быстрее. Для метаболизма покоя показатель степени при массе равен 0,75, а при нагрузках он достигает у млекопитающих даже 0,81.
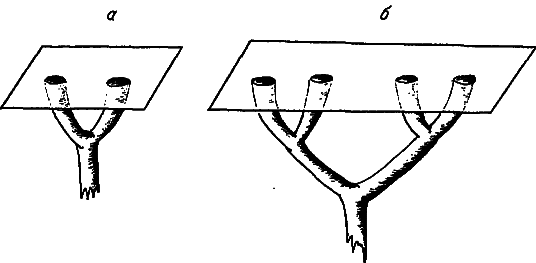
Рис. 7. Усложнение ветвления одномерных структур при увеличении размеров органов.
а — простая структура; б — сложная.
По-видимому, некоторое превышение реальной мощности метаболизма над уровнем, достаточным для снабжения энергией скелетной мускулатуры, ведет к ускорению бега животного. Скорость бега можно определить, перемножив частоту шагов на длину среднего шага. Измерения показали, что с увеличением размеров животных частота их шагов снижается существенно медленнее, чем возрастает длина шага, в итоге бег ускоряется.
Итак, увеличенные размеры дают определенные преимущества. Во-первых, без снижения скорости (и даже при ее некотором росте) бег становится более экономичным. Во-вторых, падает удельная мощность метаболизма, что фактически означает уменьшение энергетических расходов на поддержание жизни единицы биомассы. Однако эти, казалось бы, явные плюсы сменяются на минусы при беге в гору. Для поднятия единицы массы тела на одну и ту же высоту животным вне зависимости от их размеров нужно затратить одно и то же количество энергии, но удельная мощность метаболизма у мелких животных больше, поэтому при беге в гору им надо повысить эту мощность в меньшее число раз, чем крупным животным. Так, при беге по вертикали мышь увеличивает потребление кислорода на 20 %, а лошадь — на 630 %. Значит, бегать по пересеченной местности мелким животным легче.
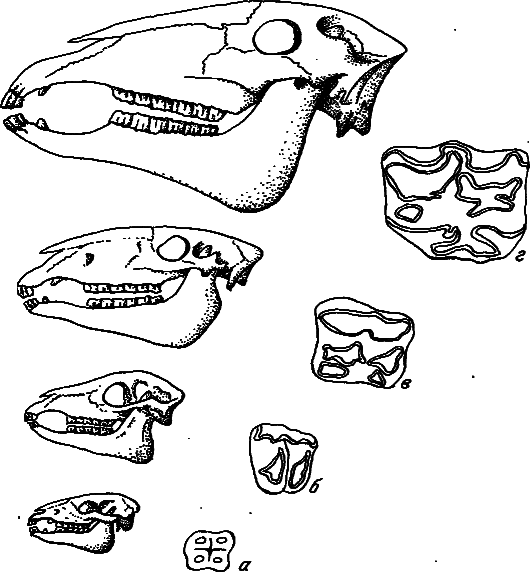
Рис. 8. Усложнение поверхности зубов у лошадей в процессе эволюции (по: [Хадорн, Венер, 1989]).
а — Eohippus (эоцен); б — Mesohippus (олигоцен); в — Merychippus (миоцен); г — Equus (плиоцен — современность).
Время жизни особи принято измерять годами, днями и т. д., хотя его можно было бы отсчитывать и числом каких-нибудь внутренних циклов — числом сердцебиений, дыхательных движений, опорожнений кишечника, делений клеток определенного типа и т. д. На выполнение каждого такого цикла требуется энергия, в связи с этим поток энергии, приходящийся на единицу массы тела (т. е. удельную мощность метаболизма р), можно выбрать в качестве меры скорости протекания всевозможных циклических процессов. Чем эта скорость выше, тем больше циклов совершается в организме за единицу астрономического времени, т. е. тем быстрее течет собственное время индивида. Темп жизни животного ускоряется с уменьшением его массы (см. (2.5)). Это означает, что мелкие животные способны реагировать на изменение окружающей среды быстрее, чем крупные. Обратная сторона медали заключается в ослаблении устойчивости к энергетическому голоду. Хорошо известно, что для крошечных землероек прекращение питания даже на 7–9 часов равносильно гибели.
Попробуем выяснить, как соотносятся между собой размеры тела и рассмотренная выше сложность морфологических структур. Сначала отметим, что увеличение органа может привести к усложнению одномерных структур, обеспечивающих связь этого органа с другими частями организма. На рис. 7 изображено, как повышается ранг ветвления сосудов, протоков, нервов и т. д.
Кроме того, есть веские основания считать, что изометрический рост ведет к непропорциональному увеличению функциональной нагрузки на многие органы. Мы уже отмечали, что при укрупнении животных их суммарный метаболизм возрастает по меньшей мере пропорционально М0,75. Однако при изометричном росте площади двухмерных структур увеличиваются существенно медленнее, следуя за М0,67. Если эти площади прямо отражают мощности соответствующих структур, то пропорциональный рост приведет к их относительному ослаблению. Отсюда следует, что функционально важные поверхности должны расти быстрее, чем М0,67, т. е. аллометрично.