Если исследовать все многообразие взаимодействий между организмом и средой, а также между различными частями одного организма, то нам откроется бесчетное число функций, подчиненных основным. Возьмем, к примеру, кости конечностей позвоночных. Мы найдем здесь и поверхности суставов, и отростки для прикрепления мышц, и отверстия для нервов и сосудов, т. е. морфологические признаки, имеющие четкую функциональную нагрузку. Распилив кость, обнаружим, что костные балки располагаются вдоль линий сжатий и растяжений, в полном соответствии с теорией сопротивления материалов. Кроме того, в полостях трубчатых костей млекопитающих мы найдем кроветворную ткань — костный мозг. А у птиц эти полости заполнены воздухом и связаны с легочными мешками; получается двойной функциональный эффект — масса тела уменьшается, а дыхание интенсифицируется.
Подобный анализ можно продолжать бесконечно, и всюду мы будем находить функциональное назначение морфологических структур. Этот поиск и обнаружение целесообразности в строении живых организмов настолько завораживают, что возникает невольная мысль о целесообразности их малейших черточек. Такая точка зрения, безусловно, относится к ненаучным, так как в реальном мире нет места для идеальных объектов. Совершенно ясно, что доведенная до предела целесообразность автоматически требует признания существования бога.
Обобщив результаты нашего функционального анализа, можно прийти к заключению, что живое вещество фактически является лишь средством для жизни как процесса, состоящего в производстве этого же вещества, т. е. цель и средство здесь удивительным образом переплетены. Получается, что для потребления энергии из окружающей среды живая система должна обладать большим набором функций и постоянно затрачивать энергию на синтез соответствующих рабочих структур. Вообще, живое вещество можно рассматривать как единый катализатор, способный сам себя синтезировать в процессе катализируемой реакции. Ясно, что масса живого вещества будет расти в геометрической прогрессии, и в той же прогрессии будет нарастать энергетический поток внутрь живой системы. Предел этому может положить только ограниченность ресурсов внешней среды.
Итак, любая живая система (будь то людское общество или природная популяция) для продления своего существования должна постоянно потреблять из окружающей среды энергию и вещество. Большая часть этого потока растрачивается на тепло и отбросы, а остальная идет на создание и поддержание рабочих структур, сохраняющих мощность энергетического потока вовнутрь системы на прежнем уровне.
Каждая такая структура выполняет свой, специфичный для нее вид работы. Мерой функциональной эффективности структуры может служить ее мощность, т. е. объем работы, выполняемой за единицу времени в режиме максимальной нагрузки. Нередко структура является необходимой, но инертной частью какого-нибудь сложного аппарата (например, локомоторного), или же ее функциональное назначение связано с пассивной защитой (например, панцирь с шипами). В этом случае строение структуры должно соответствовать максимальной нагрузке со стороны каких-то активных внешних или внутренних факторов. Способность уравновешивать подобные нагрузки также можно назвать мощностью. Это позволяет говорить о мощном костяке, мощных крыльях, мощном панцире и т. д.
Достаточно беглого взгляда на рис. 4, Б, чтобы сказать, что по мощности легких млекопитающие значительно превосходят земноводных. Выходит, что из анализа строения ряда гомологичных структур мы можем извлечь информацию об их функциональном назначении и мощности. В целом же все рабочие структуры — от молекулярных до морфологических — можно считать информационными. Выражаясь более антропоморфно, любая живая система обладает «знанием» об условиях преобразования энергии и вещества внешнего мира в собственную биомассу.
Аллометрия
Родственные виды животных обычно различаются средними размерами своих особей. Кроме того, типичные особи могут различаться по многим количественным характеристикам (параметрам), таким как величина любого органа, потребление кислорода, число митохондрий в клетке печени, средняя продолжительность жизни и многое другое. Оказалось, что для совокупности видов одного таксона величины подобных параметров (X) очень часто связаны с массой особи (М) так называемой аллометрической зависимостью
X = а*Мα, (2.2)
где а и α — аллометрические коэффициенты.
Заметим, что зависимости такого рода связывают две характеристики типичной особи каждого вида в пределах всей таксономической группы.
Известно, что при увеличении размеров шара его объем возрастает пропорционально кубу, а поверхность — только квадрату радиуса. Этот же закон обычно выполняется и при пропорциональном (изометричном) увеличении тел более сложной формы: длины всех одномерных структур возрастают пропорционально кубическому корню из массы тела, т. е. пропорционально М0,33, а площади всех поверхностей — пропорционально М0,67.
Выберем в качестве параметра одну из важнейших характеристик особи — скорость потребления ею энергии в состоянии покоя, т. е. мощность основного метаболизма (обмена веществ). Этот параметр можно оценить либо по скорости потребления кислорода, либо по скорости рассеивания тепла. Естественно, при увеличении размеров особи растет и мощность ее метаболизма (Р). Эта связь выражается эмпирическим уравнением
Р = а*М0,75. (2.3)
Здесь коэффициент а численно равен мощности основного метаболизма особи с единичной массой (скажем, с массой 1 кг). После логарифмирования уравнение (2.3) приобретает вид
ln Р = ln а + 0,75 ln М. (2.4)
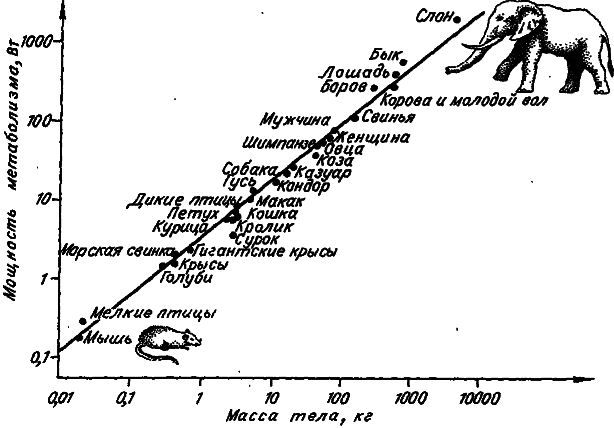
Значит, в логарифмическом масштабе мы должны иметь линейную связь между мощностью метаболизма особи и массой ее тела. Если эту связь представить графически, то все плацентарные млекопитающие (от мыши до слона) и птицы (кроме воробьиных) попадут на одну прямую, наклон которой равен 0,75 (рис. 6). Все эти животные имеют очень близкие значения коэффициента а. Для сумчатых, плацентарных млекопитающих и воробьиных птиц данный коэффициент равен соответственно 46,5; 73,3 и 129,0. Для амфибий и рептилий его значения колеблются в диапазоне от 0,8 до 7,8. Получается, что особи одной массы, но принадлежащие к разным таксонам, рассеивают в состоянии покоя разное количество энергии.
Рис. 6. Кривая «от мыши до слона». Зависимость мощности метаболизма от массы у млекопитающих и птиц (по: [Benedict, 1938]).
Разделив обе части уравнения (2.3) на М, получим связь массы тела со скоростью рассеивания энергии единицей массы животного, т. е. с удельной мощностью метаболизма (р):
р = а*М-0,25. (2.5)
Знак минус при показателе степени свидетельствует о том, что с ростом массы особи удельная мощность метаболизма падает. Эта закономерность верна для всех животных, объединенных общим значением коэффициента а. Заметим, что у млекопитающих размеры гомологичных клеток отличаются незначительно, поэтому уравнение (2.5) указывает на ослабление метаболизма средней клетки при увеличении массы животного.
Жизнь особи многообразна. Животное может и мирно отдыхать, и мчаться изо всех сил, спасаясь от смертельной опасности или преследуя свою жертву. Совершенно ясно, что в течение своей жизни организм неоднократно испытывает максимальные физические нагрузки. Главные энергетические расходы животных, безусловно, связаны с активным движением. Достаточно отметить, что при интенсивных мышечных усилиях потребление кислорода возрастает в 10–30 раз. Поэтому мы должны прежде всего рассмотреть, как изменяется режим работы мышц при увеличении размеров особи.
Как известно из механики, работа (A) равна произведению силы на путь. При сокращении мышцы на длину Δl она совершает работу в соответствии с формулой