Настоящие халикотерии, приспособившиеся к жизни в лесной чаще и питавшиеся листьями, побегами и плодами деревьев, просуществовали до середины плейстоцена, хотя, судя по относительно небольшому общему числу форм, никогда не играли ведущей роли в древних фаунах млекопитающих. Эти когтепалые, преимущественно крайне низкокоронковые животные, едва ли могли легко передвигаться по топким или даже просто мягким грунтам (сильно сплющенные с боков и заостренные дистально когтевидные копыта не адаптированы к таким условиям передвижения). Однако поскольку низкокоронковые зубы халикотериид, приспособленные к питанию мягкой и сочной растительностью, указывают, несомненно, на связь с влажным лесом, в особенностях строения их конечностей мы усматриваем адаптацию к передвижению по заваленной листвой, ветками и стволами деревьев почве, а иногда, у мелких форм, даже к лазанию по деревьям с невысокой и сильно разветвленной кроной.
Все же несмотря на то что халикотерии занимали специальную нишу, их ареал, судя по имеющимся у нас данным, резко уменьшился к концу плиоцена и в плейстоцене ограничился только некоторыми областями Азии. Это, возможно, связано как с широким распространением в конце миоцена и в начале плиоцена охотившихся на них пальцеходящих хищников, так и с заметным иссушением климата, вызвавшим сокращение влажных биотопов, представлявших местообитания этих животных. Полное исчезновение халикотериид было ускорено также инадаптивным строением их коренных зубов.
Обращаясь к бронтотериям (рис. 23), мы должны заметить, что в отличие от халикотериид эта группа непарнопалых имела свой период расцвета (средний и поздний эоцен), когда ее многочисленные представители населяли всю голарктику.
К началу олигоцена наблюдается, однако, заметный упадок в развитии этих непарнопалых, и хотя в это время некоторые формы бронтотериид достигают максимального увеличения размеров и крайней специализации признаков черепа и зубного аппарата, они, по-видимому, не переживают среднего олигоцена. Все же нельзя сказать, что существование бронтотериид резко прерывается в середине олигоцена, ибо в позднем эоцене существовало двенадцать родов бронтотериид, в ранем олигоцене их стало восемь, а в среднем сохранилось два или три рода и при этом заметно сократился их ареал (среднеолигоценовые формы известны только в Монголии).
Нам представляется вполне приемлемым объяснение причин вымирания бронтотериид, предлагаемое Осборном (1929). По мнению этого автора, бугорчато-лунчатые зубы бронтотериев, состоящие из двух обособленных бугорков на внутренней стороне верхнего коренного и двух полулунных гребней на наружной, являются инадаптивыми, так как из таких зубов не могут развиваться ни высококоронковые, ни гребенчатые зубы. За отсутствием поперечных гребней, соединяющих внутренние бугорки с наружными гребнями, увеличение высоты коронки привело бы к развитию изолированных столбиков и полулуний. В процессе эволюции у бронтотериев увеличивалась высота полулуний, а бугры оставались низкими.
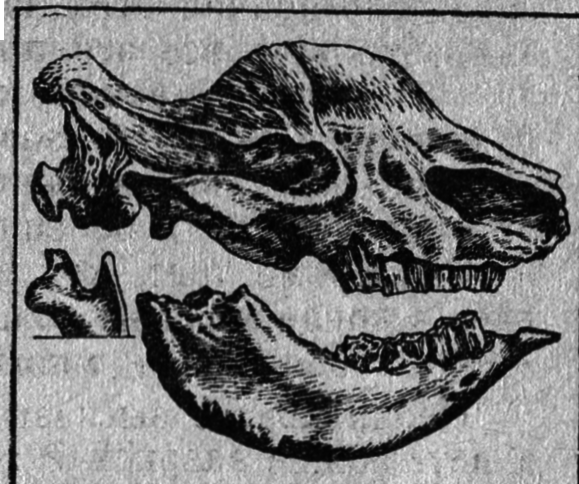
Рис. 24. Эласмотерий (кости черепа)
В результате получились наполовину гипсодонтные и наполовину брахиодонтные [2] зубы.
Такие коренные зубы у бронтотериев, естественно, не могли служить исходным типом для развития коренных зубов копытных, питавшихся относительно жесткой травянистой растительностью. Если добавить к этому массивность и явную инадаптивность строения конечностей, а также неуклюжесть животных в целом, станет очевидным, что бронтотериям было трудно выдерживать конкуренцию со стороны более совершенно организованных и более подвижных форм непарнопалых. В частности, различные группы носорогов, распространившиеся с позднего эоцена и раннего олигоцена по всей голарктике, были опасными конкурентами для бронтотериид, ставших на путь захвата относительно сухих стаций. Обладая лофодонтными зубами и относительно стройными конечностями, носороги, особенно экологически близкие к бронтотериям аминодонты и отчасти индрикотерии, могли относительно быстро вытеснить последних бронтотериид из занимаемой ими экологической области.
Халикотерии, обладавшие такими же буно-селенодонтными зубами, как и бронтотерии, просуществовали до плейстоцена только благодаря отмеченному выше своеобразию их строения, позволившему им надолго закрепить за собой специальные экологические ниши леса.
Находим небезынтересным также вопрос о вымирании эласмотериев (рис. 24), одной из наиболее любопытных групп носорогов, выработавших в процессе эволюции более совершенную, чем у лошадиных, гипсодонтность. Исчезновение эласмотериев объяснить нелегко. Эти носорогообразные были вполне приспособлены к жизни в сухих саваннах и полупустынях. Их зубы были как нельзя лучше адаптированы к питанию жесткой растительностью. Весьма крупные размеры и, вероятно, толстая, как у прочих носорогов, кожа делали их трудноуязвимыми для хищников. И тем не менее настоящие эласмотерии (род Elasmotherium), существовали недолго (с позднего плиоцена до среднего плейстоцена), а область их распространения была относительно невелика (Восточная Европа, Средняя Азия, Китай). Однако внимательное исследование признаков строения эласмотериев дает основание считать эту ветвь носорогов в некоторой степени инадаптивной.
Жизнь в открытых пространствах требует быстрой ориентации и подвижности, связанных не только с необходимостью избегать встречи с хищниками, которые не представляли, быть может, особой угрозы для эласмотериев, но и для освоения хороших пастбищ. Отлив же головного мозга эласмотериев свидетельствует об относительно низком уровне развития центральной нервной системы. У них огромные обонятельные доли были расположены перед небольшими и слабо развитыми полушариями переднего мозга. Если учесть, что другие носороги уже в миоцене достигли свойственного современным формам усложнения головного мозга или даже превзошли их в этом отношении, то такое исключительное положение эласмотерия может вызвать удивление.
Одно из возможных объяснений причины резкого отставания эласмотериев от других носорогов в развитии головного мозга заключается в следующем допущении. Относительно быстрое увеличение высоты коронки зубов давало ранним эласмотериям такие громадные преимущества перед другими носорогами и прочими копытными, захватившими степные и полупустынные стации, что эта новая обстановка не предъявляла в начале их центральной нервной системе особых требований. Это вызвало некоторый «застой» в развитии головного мозга. Впоследствии же, когда широко распространились настоящие лошади и настоящие антилопы — вероятные конкуренты плейстоценовых эласмотериев, биотическая обстановка сильно осложнилась. В условиях этой новой обстановки низкоорганизованных эласмотериев не могли уже выручать гипсодонтныё коренные, тем более что в целом животное оставалось массивным и неуклюжим, а его трехпалые конечности не были в достаточной мере приспособлены к быстрому передвижению. Наконец, вполне вероятно, что последний удар был нанесен эласмотериям похолоданием, связанным с оледенением.
Следует отметить, кроме того, что позднейший, из известных представителей эласмотериев был современником раннепалеолитического человека. Можно думать поэтому, что и человек каменного века сыграл некоторую роль в его окончательной экстерминации.
Заметим в заключение, что в раннетретичный период истории млекопитающих многие группы становились на путь скорейшего приобретения признаков специализации, обеспечивающих им относительное преуспевание. Однако, как показывает анализ этих сравнительно быстро развивающихся приспособительных особенностей палеогеновых млекопитающих, почти все они явно инадаптивного характера и ведут лишь к недолговечному господству их обладателей. Действительно, очень немногие инадаптивные ветви палеогеновых млекопитающих пережили олигоценовую эпоху и только две из них, тапиры и носороги, достигли наших дней. Все это с несомненностью свидетельствует об огромной роли инадаптивной эволюции в истории млекопитающих. По-видимому, этот способ эволюции являлся весьма важным условием вымирания млекопитающих, особенно раннетретичных групп класса Mammalia.