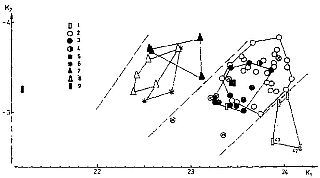
Поляки, литовцы и особенно латыши и эстонцы представлены наибольшей макросомностью: лептосомией, относительно длинными конечностями, относительной узкоплечестью и центрифугальностью. Те же свойства, исключая узкоплечесть, встречаются у русских и молдаван, а также у немцев.
Народы Кавказа, Средней Азии и Поволжья характеризуются микросомностью, относительно небольшой длиной конечностей, центрипетальностью. Континентальные монголоиды, представленные в данном случае якутами, составляют еще одну обособленную группу, в которой микросомия сочетается с лептосомностью, относительно короткими конечностями и центрипетальностью поперечного развития тела.
Русские Калининградской области ничем не отличаются от остальных групп русских и отделены от прибалтов и поляков (как и русские Ленинградской области — от эстонцев и латышей). Русские Восточно-Казахстанской и Читинской областей отличны от европейских русских, но не вливаются в общность монголоидного кластера. То же самое можно сказать и о русских Амурской области и Приморского края. Следовательно, речь идет скорее о влиянии среды, чем о генетической трансформации.
Межгрупповая вариация сочетания средних значений первых двух канонических переменных телосложения (крестиком в кружке обозначены русские азиатской части территории СССР и Калининградской области).
1 — народы Прибалтики, поляки, немцы,
2 — русские, 3 — украинцы, 4 — белорусы,
5 — молдаване, 6 — народы Поволжья, 7 — народы Закавказья,
8 — народы Средней Азии, 9 — якуты.
[Балты, славяне, прибалтийские финны. Этногенетические процессы. Рига: Зинатне, 1990.]
Геногеография Русского мира
Большой проблемой для расологических выводов являются данные геногеографических исследований, которые могут интерпретироваться самым превратным образом.
Если антропология оценила русских как высокоинтегрированый народ, то в генетике, в силу несовершенства методик, возникают соблазны представить русских как народ гетерогенный. Параметр гетерогенности по 44 классическим маркерам (иммунологические маркеры, включая группы крови AB0 и Резус, физиологические маркеры — генетически заданные особенности ощущения цвета и вкуса, генетико-биохимические маркеры — ферменты эритроцитов и сыворотки крови) оказался равным 2.00. При значительно меньшем числе изученных популяций и классических маркеров для других европейских народов, генетики решили опубликовать достаточно сомнительные материалы, в которых гетерогенность русских выглядит весьма высокой. Так, болгары, чехи, словенцы, поляки имею гетерогенность в пределах 0,2-05. Примерно в тот же диапазон укладываются шотландцы, англичане, норвежцы, шведы, датчане, немцы. Адыги, кабардинцы, черкесы, абазины, абхазы, чеченцы, ингуши, народы Дагестана имеют гетерогенность в диапазоне 04–08. Более высокий уровень гетерогенности обнаруживается у австрийцев (2,34), исландцев (1,23), итальянцев (1.71), французов (1,19), румын (1.34), грузин (1.25), осетин (2.67). Аномально высокую гетерогенность вблизи показателя 7.0 зафиксировали у коми, тофаларов, эвенков нанайцев, коряков, нивхов.
Совершенно иные результаты возникают при анализе данных по одному и тому же набору параметров и при усреднении данных по территории (то есть вычислении средней гетерогенности — типичной для народов, проживающих на данной территории). Тогда у западноевропейцев и кавказцев типичный уровень гетерогенности — 0,7–0,8; для восточноевропейцев, жителей Предуралья, Зауралья, Средней Азии и Казахстана — 1,2–1,4; для жителей Сибири и Дальнего Востока — около 4.
Этот метод вычисления и сравнения полученных данных вызывает серьезные нарекания и связан скорее с начальным этапом исследований и предварительными, весьма неточными результатами, которые могут быть пересмотрены позднее — при введении в оборот большего числа данных. Необходимо подчеркнуть, что здесь, как и всюду, генетики предпочитают исследовать сельские популяции, оставляя проблему генетического разнообразия в городах в стороне.
В целом межпопуляционная дифференциация скорее говорит об оценке уровня оседлости, чем о разнообразии типов. Ведь помимо межпопуляционного есть еще внутрипопуляционная гетерогенность. Вероятно, у русских она очень высока. Только так можно совместить данные генетических исследований с антропологией, говорящей, что размах разнообразия у русских вдвое ниже, чем у западноевропейцев. Хорошая изученность русских по классическим маркерам включает в себя также исследование периферии ареала, который дает основной вклад в межпопуляционное разнообразие. Для остальных народов исследования были более скудными и, разумеется, периферию вряд ли включали.
Собрание данных о генотипе населения Русского мира для установления родственных связей и типов этих связей без дифференциации генов по значимости создает иллюзии простых интерпретаций. Например, географические карты, составленные для главных компонент, очевидным образом отдают приоритет одной группе генов в сравнении с другими. Причем, без всякой на то «физической» причины — только из удобства расчетов. Соответствующие геногеографические карты скорее задают новые загадки, чем позволяют разгадывать те, ради которых они составлялись. К тому же надо учитывать погрешности метода, потребовавшего специальных способов обработки данных.
И все же при всей своей фрагментарности и отсутствия общей системы геногеографические исследования выявили фундаментальную закономерность: основные признаки антропологических и геногеографических карт для русских оказываются идентичными. Причем очень близкими также являются карты, полученные на основе исследования классических маркеров и ДНК-маркеров. Это дает основания полагать, что выявлены основные территориальные особенности русского генофонда. Наиболее характерная географические изменчивость генофонда русского ареала имеет широтную картину. Эта картина принципиально отличается от преимущественно долготной изменчивости у населения Восточной Европы в целом — здесь изменчивость имеет преимущественно долготную протяженность геногеографических изолиний. Данное обстоятельство говорит о собственной истории генофонда русских, отличной от истории остальных народов, живущих с ними по соседству.
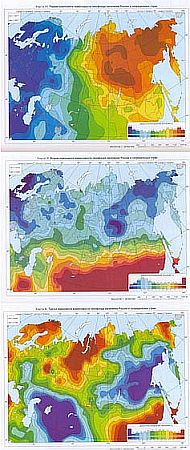
Метод главных компонент, примененный к анализу свыше 100 аллелей и гаплотипов 35 локусов полиморфных генов населения Северной Евразии, дает сложную картину пространственного распределения. Три первые главные компоненты охватывают 40 % обобщенной дисперсии частот. Первая главная компонента описывает для России и сопредельных стран 25,8 % разнообразия, вторая главная компонента — 11,9 %, третья — 6,7 %. К сожалению, нет уверенности в том, что отраженная в картах изменчивость ценнее той, которая осталась за пределами картографирования. Тем не менее, очевидно разделение Северной Евразии на генетически однородную Европу и сильно дифференцированные Сибирь и Дальний Восток, где генетики изучали преимущественно «коренное» население, исключая русских.
Первая главная компонента показывает серьезные отличия приуральского и зауральского населения от населения Восточно-европейской равнины — нарастание отличия идет почти строго с запада на восток.
В совокупности первые две главные компоненты достаточно отчетливо демонстрируют Урало-Амурский геногеографический барьер, обусловленный не только географическими и климатическими факторами, но и взаимным давлением европеоидов и монголоидов в южно-уральском узле и в Средней Азии. Третья главная компонента показывает максимум изменчивости генофонда именно в этой конкурентной зоне.
Первая, вторая и третья главные компоненты изменчивости генофонда Северной Евразии
[Генофонд и геногеография народонаселения. Том 2. Геногеографический атлас населения России и сопредельных стран. СПб.: Наука, 2003.]