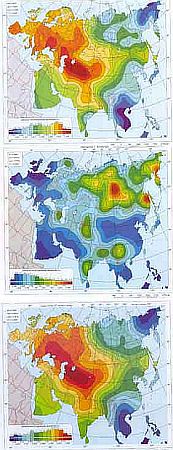
Для представления о генетическом портрете континента весьма важна картина гетерозиготности, которая является интегральным показателем микроэволюции генофонда. В данных, представленных на карте, уровень гетерозиготности исчислялся по 33 локусам. Низкая гетерозиготность наблюдается на северо-востоке Азии — у чукчей и эвенов, а также у наганасан Таймыра и эвенков Средней Сибири. Альтернативная зона минимума — в восточной части Индокитая. Максимум гетерозиготности приходится на Среднюю Азию. Средние величины наблюдаются в Передней Азии и на Аравийском полуострове.
Гетерозиготность, межпопуляционное генное разнообразие и внутрипопуляционное генное разнообразие.
[Генофонд и геногеография народонаселения. Том 2. Геногеографический атлас населения России и сопредельных стран. СПб.: Наука, 2003.]
Межпопуляционное разнообразие оценивалось на уровне самой мелкой популяционной структуры. Область высоких значений Средней Сибири окружена областями низкого уровня различий между соседними популяциями. Альтернативные максимумы — в нижнем течении Амура и на Колыме.
Внутрипопуляционное разнообразие составляет основную долю тотального разнообразия (карта гетерозиготности), отчего соответствующие карты очень сходны с картами общего разнообразия. Область максимума занимает обширную область от Памира до северо-восточных берегов Каспия. Обширная полоса средних значений захватывает Западную Сибирь и Урал. Минимум приходится на территорию северной части Дальнего Востока и Индо-Китай.
Генетические маркеры позволяют выделять геногеографические зоны — предположительные центры этногенеза. Целый ряд регионов Евразии отличается особой частотой целого ряда маркеров. Огромный материал геногеографических исследований показывает как периферийные, так и континентальные локализации потенциальных центров этногенеза:
Периферийные: Аравийский п-ов, юг Индии, восток Индии, Пиренейский п-ов, низовья Амура, Индокитай, Таймыр, север Якутии, Восточный Китай, Северное Причерноморье, Северный Кавказ, юг Камчатки, север Скандинавии и др.
Континентальные: центр Монголии, Прибайкалье, Приаралье, Казахский мелкосопочник, Памир, срединное течение Иртыша, Амурско-зейская равнина, бассейне Хатанги, бассейн реки Ижмы, Северное Причерноморье, район Нижнеенисейской возвышенности, Чукотское нагорье, локус в китайских Гималаях, район Череповца и Ярославля, зона Непала, зона Ладожского озера и др.
Множественные континентальные локусы в Сибири и на Дальнем Востоке относятся к малым народам и малосущественны для генетических перспектив континента. Большинство малых локусов или малонаселенных территорий лишь в перспективе может дать вклад в евразийский генотип. Куда существенней значительные континентальные пространства, отмеченные особенностями в связи с остальными евразийскими территориями. Для нас важно выделить присутствие генных аномалий в прикаспийском и приаральском пространстве, откуда, как можно предположить, происходили основные миграционные процессы, определившие генетический и антропологический портрет Европы и Русского мира.
Генофонд и расово-диагностические признаки
Группы крови человека системы АВ0 обозначаются символами: 0(I), А(II), В(III), АВ(IV). Они кодируются тремя аллельными генами: 0, А и В, первый из которых рецессивный, остальные — кодоминантные. Наследование дает сочетания АА, А0, ВВ, В0, АВ, 00, которые воплощаются в фенотипах А, В, АВ и 0. Сегодня известны две модификации гена А1 и А2, а также множество прочих модификаций различной специфичности.
По географии частотности распространения той или иной группы крови, а также по географии «кровного разнообразия» можно судить как о различиях между человеческими расами, так и о возможных древних миграциях. При низкой дифференциации этносов по крови, на больших пространствах дифференциация становится заметной.
Аллель А системы АВ0 считается слабодифференцирующим человеческие расы и этносы. Вместе с тем, она почти не встречается в Новом Свете у коренных народов (в среднем 0,033), низкий показатель частоты присутствует в Африке (0,160), несколько выше в Меланезии (0.190), Восточной Азии (0.193), в Австралии (0.192) и Индии (0.195). Континентальный максимум, незначительно отличающийся от средней нормы, наблюдается в Европе и Западной Азии (0.230) и Океании (0.241). Минимум частоты наблюдается у многих сибирских популяций — у бурят (0.098), восточных эвенков (0.018), манси (0.089), лесных ненцев (0.083).
В Средней Азии показатель особенно высок у памирских рушанцев (0.636), в Сибири — у западных эвенков (0.559), на Кавказе — у армян и аварцев (0.401) и лакцев (0.410). Минимальный показатель наблюдается у таджиков (0.086). В Европе выше среднего показатель у башкир (0.379), великорусов (0.428), белорусов (0.430), саамов (0.371), ниже среднего у карагашей, калмыков и караимов (0.186, 0.184, 0.161).
Межпопуляционное разнообразие по аллелю А не демонстрирует заметных аномалий в европейских этногеографических зонах. В сравнении с Европой, показатель в Сибири и на Дальнем Востоке несколько понижен. В целом в Северной Евразии частота аллеля повышается с запада на восток.
Евразийское распределение частоты аллеля А указывает на максимумы в периферийных зонах (север Скандинавии и запад Пиренейского полуострова). Средние значения охватывают почти всю Европу, Переднюю Азию и Приаралье, образуя общее пространство, разорванное лишь на Северном Кавказе и в Северном Прикаспии. Картина предполагает возможность миграционного потока ариев из Приаралья на восток и северо-восток. Северокавказский разрыв может быть обусловлен инорасовой волной миграции с востока и северо-востока.
Частота А1 на севере евразийского пространства максимальна у русских Тверской области (0.381), минимальна — в ненцев (0, 059). Не считая малолюдных зон севера Сибири (с очевидно низкой достоверностью измерений), на евразийской карте частоты А1 видны три континентальных максимума — в Центральной России, в южном Прикаспии и в континентальном Китае. Если прикаспийскую зону можно считать периферийной (зоной, куда были оттеснены мигранты), то два оставшиеся центра следует считать источниками этногенеза — в первом случае славянского, во втором — китайского.
Средняя частота аллеля А2 в Европе составляет 0.100, в Африке и Западной Азии снижается до 0.05, а в других регионах практически отсутствует. Частота А2 высока лишь у алеутов, финнов и саамов (0, 344, 0.213, 0.310). Высокие значения (выше 0.200) характерны для русских, марийцев, белорусов. У многих народов Сибири и Дальнего Востока, а также у казахов и узбеков частота близка к нулю. Локальные максимумы наблюдаются у саамов Норвегии (0.235) и чеченцев пос. Ведено (0.142)
Распределение аллеля А2 на карте Евразии дает двоякую картину: подтверждает периферийность севера Скандинавии и демонстрирует ярко выраженный локус этногенеза в южном Китае. В пространственных распределениях демонстрируется общность юго-западной Азии и Европы и противоположность этой геногеографической зоны остальной части Азии.
Аллель В считается слабодифференцирующим по этнической принадлежности. Наибольшая частота наблюдается в Индийском регионе (0.240), наименьшая — у коренных американцев (0.015) и в Австралии (0.021). В Европе этот показатель составляет в среднем 0.184, на Кавказе — 0.130. Высокие частоты встречаются у народов Сибири и Дальнего Востока.
В европейском регионе по этому показателю выделяются марийцы (0.398), на Кавказе — дагестанцы (0.345), в Средней Азии — узбеки (0.383). Низкие частоты отмечены у русских (0.082), белорусов (0.071), украинцев (0.043), на Кавказе — у армян (0.081) и грузин (0.033), в Азии — у чукчей (0.083), шорцев (0.063), алтайцев (0.043), эвенков (0.020).
Аллель В на карте Евразии демонстрирует картину, во многом обратную картине распределения аллеля А2. При этом азиатское пространство заполнено множеством локальных максимумов распределения. Крупные зоны в Иране, Индии, Китае и Приморье соседствуют с множеством более компактных зон максимума частоты аллеля В. Возможно, это свидетельство более поздних процессов этногенеза, характерных для азиатских народов.