Глава VII
У истоков генетики
Слабое место в теории Дарвина
Процесс передачи потомству родительских наследственных факторов долгое время оставался совершенно непонятным. Открытие в конце XVII в. сперматозоида вызвало целую дискуссию. Одни утверждали, что будущий зародыш целиком заключен в яйцеклетке и оплодотворение является лишь толчком к развитию. Другие настаивали на том, что будущий зародыш помещается в сперматозоиде, а яйцеклетка лишь обеспечивает его питание. Спорщики сходились на том, что носителем наследственных факторов является одна родительская особь.
Только в середине XVIII в. в результате наблюдений над детьми от смешанных браков у человека и исследования экстерьера мулов было установлено, что признаки наследуются от обоих родителей. Пьер Луи Моро Мопертюи (1698–1759) выдвинул теорию, согласно которой наследственные признаки у потомков формируются и определяются «семенными частицами» обоих родителей.
Даже в XIX в. еще не было правильного представления о наследственном механизме. Именно поэтому эволюционная теория так часто применялась неудачно. Спенсер считал возможным быстрое изменение человеческого поведения: Гальтону казалось весьма несложным усовершенствовать человеческую расу с помощью подбора производителей при размножении. Подобные воззрения биологов объяснялись, по сути дела, недостатком знаний о природе наследственного механизма. Это было самое слабое звено в теории Дарвина. Дарвин предполагал, что среди молодых особей любого вида наблюдается непрерывная случайная изменчивость, причем определенные изменения делают животных более приспособленными к окружающей среде. Так, жирафу тем легче прокормиться, чем длиннее у него шея.
Но где гарантия, что этот признак повторится у потомства? Вряд ли жираф выискивал особо длинношеего супруга, гораздо вероятнее, что ему пришлось встретиться с короткошеим. При скрещивании крайних вариантов происходит смешение признаков, так что у потомства длинношеего и короткошеего жирафов шея будет средней длины, — к такому убеждению привел Дарвина весь его опыт по разведению животных.
Иными словами, в результате случайного спаривания все полезные и приспособительные признаки, которые проявляются при случайной изменчивости, усредняются. Раз естественному отбору нечего «отбирать», то и эволюционных изменений происходить не будет.
Попытки биологов объяснить это явление оказались безуспешными. Немецкий ботаник Карл Вильгельм Негели (1817–1891) прекрасно сознавал, как сложно найти четкое обоснование «усреднения» и его последствий. Он предположил, что эволюционные изменения направляются в определенную сторону каким-то внутренним толчком.
Так, судя по палеонтологическим данным, предками лошадей были животные величиной с собаку, имевшие по четыре копытца на каждой стопе. С течением времени эти животные становились крупнее и утрачивали одно копытце за другим, пока не превратились в современную крупную однокопытную лошадь. Негели предполагал, что побудительным фактором этого процесса было действие некой внутренней силы. Если бы она действовала и дальше, лошади могли бы стать слишком крупными и неуклюжими и, следовательно, беззащитными против врагов; им грозило бы постепенное вымирание.
Эта теория носит название теории ортогенеза. Современные биологи ее отвергают. Как мы увидим дальше, приверженность Негели к этой теории неожиданно принесла скверные плоды.
Горох Менделя
Проблема была решена благодаря трудам чешского натуралиста Грегора Иоганна Менделя (1822–1884). Мендель занимался и математикой и ботаникой. Начиная с 1856 г. он в течение девяти лет изучал наследственные признаки гороха, используя статистические приемы обработки результатов.
Ученому нужна была полная уверенность, что наследуются признаки только одного родителя, поэтому он очень тщательно проводил самоопыление различных растений, аккуратно собирал с каждого самоопыленного растения семена, отдельно высаживал их и изучал новое поколение.
В результате этих опытов Мендель обнаружил, что горошины от карликовых растений и в первом и в последующих поколениях дают только карликовые растения. Таким образом, карликовые растения чистосортны.
Высокорослые растения вели себя по-разному. Некоторые (более трети) оказались чистосортными и поколение за поколением давали высокие растения. Часть остальных семян давала высокие, а часть — карликовые растения, причем высокорослых всегда получалось в два раза больше, чем карликовых. Очевидно, имелось два типа высоких растений — чистосортные и нечистосортные.
Мендель пошел дальше. Он скрестил карликовые растения с чистосортными высокорослыми и обнаружил, что из каждого гибридного семени вырастает высокое растение. Казалось бы, признаки карликовости исчезли.
Произведя затем самоопыление каждого гибридного растения, Мендель изучил полученные семена. Все гибридные растения оказались нечистосортными. Около одной четверти горошин дали карликовые растения, одна четверть — чистосортные высокорослые, а оставшаяся половина — нечистосортные высокорослые растения.
Мендель предположил, что каждое растение гороха несет два фактора, определяющих какой-либо признак, в данном случае — высоту. Один фактор содержится в пыльце, другой — в семяпочках. После оплодотворения новое поколение содержит уже оба фактора (по одному от каждого родителя, если произошло скрещивание двух растений). У карликовых растений имеются только факторы «карликовости»; при комбинации их путем перекрестного опыления или самоопыления получаются только карликовые растения. Чистосортные высокорослые растения содержат только факторы «высокорослости», и их комбинации дают только высокие растения.
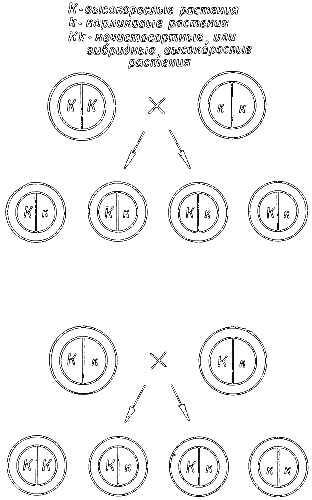
Рис. 3. Схема, объясняющая опыты Менделя по скрещиванию высокорослых и карликовых растений гороха.
Вверху — скрещивание чистосортного высокорослого растения с карликовым, которое дает гибридные, или нечистосортные, высокорослые растения. Внизу — скрещивание между собой гибридных растений первого поколения, которое дает чистосортные высокорослые растения, гибридные высокорослые растения и карликовые растения в отношении 1:2:1. Фактор карликовости обозначается буквой к, так как он рецессивен, а соответственный ему доминантный фактор высокорослости — К.
Когда чистосортные высокие растения скрещиваются с карликовыми, факторы «высокорослости» и «карликовости» комбинируются и следующее поколение оказывается гибридным. Все гибриды будут высокими, так как фактор «высокорослости» является доминантным, маскирующим действие фактора «карликовости». Последний, однако, не исчезает, а сохраняется.
Такие гибриды не будут вести себя как чистосортные, потому что содержат оба фактора, комбинации которых определяет чистая случайность. Фактор «высокорослости» может комбинироваться с другим фактором «высокорослости», тогда появится чистосортное высокорослое растение. Так произойдет в одной четверти случаев. В другой четверти фактор «карликовости» в комбинации с другим фактором «карликовости» дает карликовое растение. В оставшейся половине фактор «высокорослости» сочетается с фактором «карликовости» или фактор «карликовости» — с фактором «высокорослости»; при этом получаются высокорослые нечистосортные растения.
Мендель показал, что наследование любого признака можно объяснить таким же образом, причем для всех изученных им признаков скрещивание двух крайностей не приводило к смешению наследственности — каждый вариант сохранялся без изменения; если он исчезал в одном поколении, то появлялся в следующем.
Все это имело немаловажное значение для эволюционной теории (хотя сам Мендель никогда не думал о приложении к ней своих идей), так как позволяло сделать следующий вывод. Случайные вариации, в течение какого-то промежутка времени возникающие внутри вида, в конечном счете не усредняются, а исчезают и проявляются вновь, пока их в полной мере не использует естественный отбор.