Важнейшую роль в артикуляции звуков играет ротовая полость. Насколько можно судить по измерениям, проводившимся на ископаемых черепах, здесь также нет сколько-нибудь заметных различий между неандертальцами и людьми современного физического типа. И у тех, и у других ротовая полость устроена примерно одинаково и даёт, в общем, одинаковые возможности для произнесения различных звуков[189].
Одно время была надежда, что в качестве индикатора наличия речи удастся использовать толщину канала подъязычного нерва, находящегося в нижней части затылочной кости между яремным отростком и мыщелком и контролирующего движения почти всех мускулов языка. Поскольку можно ожидать, что интенсивность таких движений возрастала в процессе развития речи, было высказано предположение, что и сам нерв и, соответственно, канал должны были увеличиваться, а значит, размер последнего мог бы служить показателем речевых возможностей ископаемых видов. Первое сравнение площади сечения канала шимпанзе (обыкновенных и карликовых), горилл и ряда гоминид (включая современных людей) как будто подтвердило жизнеспособность этой гипотезы. Получалось, что если у австралопитеков рассматриваемая величина ещё не выходит за рамки вариабельности, свойственные человекообразным обезьянам, то у гомо гейдельбергенсис и классических неандертальцев она, наоборот, достигает тех же значений, что и у ныне живущих людей[190]. Уже через год, однако, как исходные постулаты этого исследования, так и его выводы оказались под вопросом. В частности, было показано, что у людей и шимпанзе толщина канала подъязычного нерва в значительной степени перекрывается[191]. Таким образом, к сожалению, от использования этого признака в предложенном качестве пришлось отказаться.
Зато другой похожий признак, кажется, выдержал проверку и работает вполне успешно. Это размер спинномозгового канала грудных позвонков. Для быстрой членораздельной речи необходим точный контроль дыхания. Одним из условий такого контроля у людей является рост числа тел нервных клеток в спинном мозге грудных позвонков, что имеет следствием расширение позвоночного канала грудного отдела по сравнению с другими приматами. Изучение ископаемых материалов (пока, к сожалению, очень скудных) показало, что у гомо эректус, по крайней мере ранних (тех, кого называют ещё гомо эргастер), такого расширения ещё не произошло — они в этом отношении ближе к обезьянам и австралопитекам, чем к современным людям. А вот неандертальцы, напротив, по ширине канала грудных позвонков практически не отличаются от гомо сапиенс (рис. 7.5)[192].
Наконец, детальное изучение анатомического строения наружного слухового прохода и среднего уха на височных костях пренеандертальцев из Симы де лос Уэсос показало, что на этой стадии ухо безусловно обладало теми акустическими свойствами, которые необходимы для тонкого различения звуков человеческого голоса и адекватного восприятия речи[193].
Таким образом, получается, что те анатомические органы неандертальцев, которые связаны с артикуляцией и восприятием звуков и о строении которых у нас есть более или менее достоверная информация, практически ничем не отличались от аналогичных органов гомо сапиенс. Мозг у людей обоих этих видов тоже очень похож, и если у одного из них он достаточно развит, чтобы позволить создавать и использовать для обмена информацией системы искусственных знаков, то было бы нелогично не допустить того же самого и для другого. Наконец, единственный ген, о котором сейчас более или менее достоверно известно, что он влияет на речевую деятельность, также оказался у двух видов совершенно одинаково построен, одинаковым образом отличаясь от такого же гена шимпанзе. Этот ген обозначается как FOXP2, его дезактивация имеет следствием серьёзные нарушения в работе артикуляционного аппарата. После выяснения его важности для развития речи было высказано предположение, что эту функцию, а также и специфическую — не такую, как у шимпанзе, — структуру FOXP2 приобрел сравнительно недавно и что и то, и другое свойственно только гомо сапиенс. Вскоре, однако, удалось выделить соответствующий участок ДНК из кости неандертальца, и выяснилось, что этот ген отличается от аналогичного гена шимпанзе точно по тем же двум нуклеотидным позициям, что и ген современного человека. Стало быть, мутации, приведшие к этим изменениям, должны были произойти как минимум ещё у гомо гейдельбергенсис (общего предка неандертальцев и гомо сапиенс), а возможно, и на более ранней стадии эволюции[194].
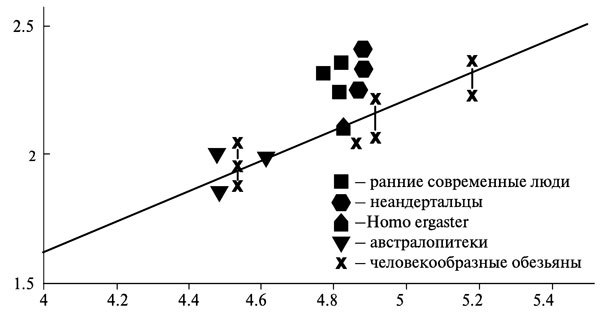
Рис. 7.5. Отношение log(10) диаметра спинномозгового канала грудных позвонков (мм) к log(10) веса тела (г) у разных групп гоминид (источник: MacLarnon and Hewitt 2004)
Из всего этого можно сделать только один вывод: потенциально неандерталец был способен к языку вообще и членораздельной речи в частности в той же или почти в той же мере, что и гомо сапиенс. Пользовался ли он этой способностью — другой вопрос, на который нет и, возможно, никогда не будет точного ответа. Однако, коль уж на то пошло, этот же вопрос можно задать и по отношению к древним представителям нашего собственного вида, жившим одновременно с неандертальцами и даже после них — и ответа тоже не будет.
Строго говоря, даже живопись и скульптура верхнего палеолита, сколь бы они нас ни впечатляли своей выразительностью, сами по себе всё же не доказывают, что их творцы разговаривали друг с другом или вообще пользовались каким бы то ни было языком. Напротив, натурализм, которым славится палеолитическая живопись, при желании можно даже интерпретировать как свидетельство интеллектуальной и языковой ограниченности её создателей. Именно такую гипотезу выдвинул несколько лет назад американский психолог Н. Хамфри.
Хамфри привел весьма остроумную аргументацию в пользу того, что общепризнанные шедевры палеолитического искусства, такие, как росписи в пещерах Шовэ и Ляско, нельзя считать доказательством наличия у их создателей развитого мышления и языка. Более того, предположил он, эти росписи могут свидетельствовать скорее об обратном, то есть об интеллектуальной неполноценности первых художников. Отправной точкой для рассуждений Хамфри послужил анализ опубликованных около четверти века назад рисунков английской девочки Нади, страдавшей аутизмом. Несмотря на то, что физически, психически и интеллектуально Надя значительно отставала от сверстников, была неспособна к сколько-нибудь нормальному общению и совсем не могла разговаривать (первые несколько слов она освоила, когда ей было шесть лет), рисунки, созданные ею в возрасте от трёх до шести лет, обладают рядом замечательных качеств. Для многих из них характерно правильное построение перспективы и абсолютная реалистичность изображений, что особенно интересно, если учесть, что рисованию Надю никто не учил и что рисункам нормально развивающихся детей того же возраста оба этих качества совершенно не свойственны. Своей натуралистичностью, а зачастую и сюжетами многие рисунки Нади удивительно напоминают широко известные произведения пещерной живописи, например, бизона из Шовэ или лошадей из этой же пещеры, а также из Ляско. По мысли Хамфри, поддержанной и некоторыми из участников обсуждения его статьи, именно отсутствие обычных для современных людей умственных способностей и, прежде всего, отсутствие языка может объяснить необычайную реалистичность Надиных рисунков. Ведь её восприятие животных или людей не было опосредовано символами, обозначающими классы объектов, и она, в отличие от обычных детей, изображала непосредственно своё впечатление от лошади или коровы, а не соответствующую абстрактную идею, т. е. между изображаемым и изображением в данном случае ничто не стояло. Предполагается, что тем же обстоятельством, то есть отсутствием абстрактных понятий, обозначающих классы объектов, может объясняться и натурализм палеолитической живописи, причём в этом случае становится понятным и переход от реалистических изображений к схематическим, наблюдаемый в конце верхнего палеолита: он связывается с появлением развитого языка и утратой непосредственности восприятия[195].