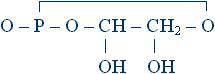Для экспериментальной оценки активности препаратов ангиотензина обычно приготавливают суспензию клеток-мишеней, добавляют в нее испытуемый препарат и следят за выделением клетками в окружающую среду альдостерона. Клетки начинают секретировать альдостерон уже в присутствии 10–10...10–9 моля ангиотензина.
До какой степени это низкая концентрация? В одном кубическом сантиметре раствора концентрации 1 моль на литр содержится 6·1020 молекул. Таким образом, в кубическом сантиметре испытанного нами раствора находится 6·1010 молекул. 60 миллиардов, не так уж как будто и мало. Правда, и клетки-мишени невелики, их линейные размеры – около микрометра, то есть 10–4 сантиметра; соответственно объем одной клетки – порядка 10–12 кубического сантиметра.
Предположим, что клетки занимают один процент инкубационной среды по объему, то есть в 1 кубическом сантиметре их окажется 1010 штук. Тем самым на каждую клетку приходится шесть молекул ангиотензина.
Приведенный расчет небезупречен; надо, скажем, считаться с тем, что в суспензии окажутся не отдельные клетки, а агрегаты клеток, что доля собственно секретирующих клеток составит не один процент объема среды, а меньше (или больше – несущественно). Во-первых, более сложные и более точные расчеты, выполненные для ряда гормонов, показали, что для развития специфической реакции клетки-мишени часто достаточно, чтобы с ее рецепторами связалось всего несколько молекул биорегулятора (по некоторым расчетам – вообще одна), во-вторых – если секреция альдостерона «защищается» не шестью, а шестьюдесятью или даже шестьюстами молекулами ангиотензина – это тоже достойно удивления, если сравнить размеры клетки и молекулы.
Опять же, в интуитивном нашем представлении и та и другая очень малы, так что не мешает сопоставить еще несколько цифр. Молекулы биорегуляторов имеют размер от нескольких десятых нанометра (скажем, адреналин) до нескольких нанометров (белковые гормоны). Размеры большинства животных клеток – порядка микрометра или нескольких микрометров. Различие в линейных размерах тем самым – 103...104. Иными словами, молекулы многих биорегуляторов рядом со своими клетками-мишенями должны выглядеть так же, как сами клетки рядом, например, с ириской или как зернышко пшена рядом с БелАЗом. Впрочем, для различных молекул и различных клеток эти оценки могут изменяться примерно на порядок в ту или иную сторону, так что это окажется зернышко уже не пшена, а гречки или мака – суть вывода не меняется: молекулы все же очень малы по сравнению с клетками, и кажется удивительным, что «посадка» нескольких ничтожных частичек на поверхность такой махины вызывает развитие в ней каких-то бурных процессов.
Ясно, что внутри клетки должны существовать какие-то системы, многократно усиливающие эффект взаимодействия молекулы гормона с расположенным на ее поверхности рецептором.
Один из самых универсальных механизмов подобного усиления открыт Э. Сэзерлендом в 1960 году. Сэзерленд исследовал действие упоминавшегося уже гормона адреналина на клетки печени, в которых он вызывает расщепление гликогена (крахмалоподобного запасного вещества) с образованием глюкозо-1-фосфата. Эта реакция катализируется ферментом гликоген-фосфорилазой, активность которой в клетках печени резко возрастает под действием адреналина. Почему?
Сам адреналин внутрь клетки не проникает, он лишь связывается с рецепторами на ее поверхности. Рецептор же адреналина образует комплекс с молекулой фермента аденилатциклазы, причем этот комплекс проходит через мембрану насквозь, так что с внутренней стороны мембраны в цитоплазму выступает активный центр фермента. Организация комплекса такова, что при связывании рецептором молекулы адреналина активизируется аденилатциклаза; детали механизма активации пока неизвестны. Субстратом аденилатциклазы является АТФ (аденозин-3-фосфат). АТФ – важнейший участник процессов превращения энергии в клетке. К молекуле аденозина (об этом соединении уже была речь выше, при обсуждении структуры нуклеиновых кислот) присоединена цепочка из трех остатков фосфорной кислоты. Аденилатциклаза отщепляет от АТФ два фосфатных остатка, а третий соединяет с остатком рибозы второй валентной связью, так что образуется цикл:
Это соединение называется циклическим аденозинмонофосфатом, или цикло-АМФ.
Цикло-АМФ выполняет функцию внутриклеточного биорегулятора (как оказалось впоследствии, не только в рассматриваемой реакции клеток печени на адреналин, но и во многих других реакциях, индуцированных гормонами). Внутриклеточным рецептором цикло-АМФ является неактивная форма фермента протеинкиназы. Происходит это следующим образом. Неактивная форма протеинкиназы – это комплекс, образованный четырьмя белковыми молекулами двух типов. Одна пара представляет собой собственно ферменты, другая – регуляторные субъединицы. Собственно, их регуляторная функция заключается в том, что, образуя с ферментными субъединицами описываемый комплекс, они лишают их каталитической активности.
Именно на поверхности регуляторных субъединиц находятся участки связывания цикло-АМФ, по два на каждой: посадка на них четырех молекул цикло-АМФ приводит к тому, что комплекс становится непрочным – от него отделяются обе ферментативные субъединицы. И в этом случае неизвестны тонкие молекулярные подробности механизмов, лежащих в основе этого эффекта, но существуют весьма надежные экспериментальные доказательства того, что в принципе все происходит именно таким образом.
Каталитически активные молекулы протеинкиназы, образовавшиеся благодаря действию цикло-АМФ, в свою очередь, активизируют фермент под названием киназа фосфорилазы. На поверхности его молекулы имеется два остатка серина; при участии протеинкиназы гидроксильные группы этих остатков фосфорилируются, необходимый для этого остаток фосфорной кислоты отщепляется от молекулы АТФ. Фосфорилированная молекула обретает ферментативную активность.
Совершенно аналогичным образом – фосфорилированием двух гидроксильных групп остатков серина – киназа фосфорилазы активирует упомянутую гликоген-фосфорилазу, которая наконец принимается за дело, начинает расщеплять гликоген.
Не слишком ли много здесь промежуточных звеньев? Рационализаторский зуд, не чуждый, по-видимому, никому из нас, подсказывает немного более простое решение: пусть с рецептором адреналина будет связана не аденилатциклаза, а сразу гликоген-фосфорилаза, которая и активизировалась бы при посадке на рецептор молекулы гормона.
Надо сказать, что подобный ход рационализаторской мысли очень распространен. Знакомясь первый раз с каким-то устройством или механизмом, мы обычно обнаруживаем в нем множество совершенно бесполезных узлов или бессмысленно усложненных элементов. Наиболее решительные принимаются тут же устранять эти очевидные просчеты конструкторов. Рассказывал мне один пожилой инженер-дорожник, как в довоенное еще время впервые появились у них грейдеры. Это были прицепные машины; толщина запорного болта в прицепном устройстве была выбрана таким образом, что при возникновении усилий, угрожающих деформациями рамы грейдера (например, если на пути встретился большой валун), болт срезался. В предвидении таких случаев завод-изготовитель прилагал к каждой машине ящик запасных болтов.
Работники же, обслуживающие этот грейдер, видели причину частых остановок просто в несовершенстве конструкции прицепного устройства; кляня на чем свет стоит бестолковых инженеров, они изготовили собственное – солидное и надежное. Через несколько дней грейдер был сдан в металлолом.
Хотя мы и говорим все чаще о клеточной инженерии, но исправлять по-своему структуру процессов, протекающих в клетке, по-настоящему еще все же не умеем. В каком-то отношении это и неплохо, ибо чаще всего нас постигла бы судьба горе-рационализаторов прицепного устройства грейдера.
Вот и высказанное выше предложение – пусть адренорецептор активирует непосредственно гликоген-фосфорилазу, а все промежуточные звенья – выкинуть. Не так все, оказывается, просто. Рецепторов адреналина на одной клетке не так уж много. Точные цифры неизвестны, но, скорее всего – сотни (от силы – тысячи, но вряд ли). Сотня молекул фермента, да еще связанного с мембраной (то есть громадные молекулы гликогена должны сами диффундировать к ним), не обеспечит должной скорости расщепления. Как же сделать, чтобы одна молекула адреналина, связавшаяся с рецептором, активировала не одну, а гораздо большее количество молекул фермента-исполнителя?