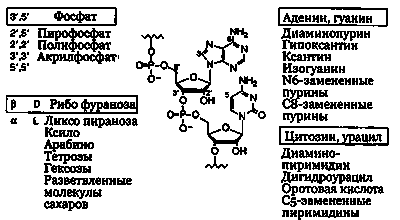
Рис. 30.5. Полимеры, сформированные фосфодиэфирными связями (содержащими фосфор и кислород) между 5'- и 3'-углеродами β-D-нуклеотидов. Нуклеозиды образованы из оснований аденина, гуанина цитозина, урацила и четырехуглеродной циклической формы рибозы в D-ориентации. Перепечатано с разрешения Macmillan Publishers Ltd; G. F. Joyce: The Antiquity ofRNAbased evolution. Nature 418:214, copyright (2002).
Еще одна трудность полимеризации рибонуклеотидов состоит в том, что в смеси мономеров могут происходить различные реакции. Чтобы сформировать функциональный полимер, фосфоризирующие соединения должны образовываться между 5'- и 3'-углеродами соседних нуклеотидов. Но кольцо рибозы имеет реакционноспособные группы ОН у углеродов в положениях 5', 3' и 2. В добиологических условиях между всеми этими группами могли протекать реакции, и в результате внутримолекулярных реакций между ОН-группами в положениях 2' и 3' могли формироваться циклические соединения. Более того, молекулы фосфатов могли создать разные полифосфатные связи между разными углеродами. Все эти разнообразные связи могли привести в тупик дальнейшую полимеризацию.
По мнению Джеральда Джойса (Исследовательский институт Скрипса, Ла-Хойя, Калифорния), ведущего специалиста по добиологической химии РНК, отсутствие специфичности является основной проблемой добиологических реакций. Спонтанные реакции, начавшиеся с синильной кислоты или с цианистого ацетилена, цианита и мочевины, могут дать разные аналоги оснований. Но из всех них только пурины аденин и гуанин, а также пиримидины цитозин и урацил природа использовала для формирования функциональных нуклеозидов. В составе нуклеозидов в добиологических условиях существующие основания могли быть связаны с составляющими рибозы, причем с одинаковым успехом как в а-, так и в р-конфигурациях, а фураноза (четырехуглеродное кольцо) рибозы могла сформироваться как в L-, так и в D-изоформах (лево- и правовращающих плоскость поляризации света, как описано в главе 28). Сахар рибоза также мог сформироваться в виде пятиуглеродного кольца (пираноза) путем соединения 5'- и 1'-углеродов. Добиологические реакции полимеризации между всеми различными аналогами и изоформами нуклеотидов могли привести к большому разнообразию фосфатных соединений разных атомов углерода рибозы. В целом эти реакции могли легко использовать разные варианты пуринов и пиримидинов, связываясь с которыми разные производные разных циклических сахаров формируют L- и D-конфигурации. Эти совершенно случайные аналоги нуклеозидов могут затем фосфорилироваться на разных позициях углерода, и потом опять случайно фосфорилированные аналоги нуклеотидов могли связаться друг с другом разными способами, что показано на рис. 30.5. Ни один из этих альтернативных вариантов не производит функциональноактивные РНК-полимеры.
Только правильно сформированные и полимеризированные нуклеотиды могут быть функциональными шаблонами для репликации через комплементарное спаривание оснований. Мы не понимаем, как жизнь при отсутствии каких-либо ферментных реакций отбора выбирала как раз нужные нуклеотидные компоненты и их специфические изоформы и как она контролировала формирование фосфоризующих связей только между 5'- и 3'-углеродами нуклеотидов.
Следующей проблемой при сборке длинных РНК-полимеров является их врожденная неустойчивость. РНК-полимеры легко делятся на части гидролизом, и их функциональная последовательность может быть утрачена из-за многочисленных ошибок копирования или мутаций. При рассмотрении всех этих химических обстоятельств кажется, что полный каскад реакций для формирования функциональных полинуклеотидов (включая синтез нуклеозидных оснований и рибозы, сборку нуклеозидов, их фосфорилирование и активацию и, наконец, полимеризацию и стабилизацию полимеров) был трудноосуществим в добиологических условиях. Эти процессы выглядят настолько непохожими, что возникло предположение, что РНК-миру предшествовали какие-то другие механизмы хранения и переноса информации, которые затем «управляли» (или обеспечивали катализаторами) миром РНК-оснований. Но трудно объяснить, как мог произойти переход от более примитивной генетической системы к РНК.
Разумеется, можно предположить, что мы пока еще не нашли таких условий, химических способов и факторов отбора, которые могли бы сделать возможной добиологическую химию и эволюцию. Новый многообещающий способ добиологического синтеза нуклеотидов — прямо из формамида — сейчас исследуется в лабораториях Р. Саладино (R. Saladino) и Э. Димауро (E.DiMauro) в университетах Тусции и Рима (Италия).
Формирование генетического кода.
Хотя молекулы РНК могут стимулировать некоторые типы химических реакций, теперь мы знаем, что белковые катализаторы значительно превосходят их по универсальности и эффективности. Поэтому изобретение генетически закодированного синтеза белков дает огромное преимущество для развития жизни. Это делает возможным появление и эволюцию основанных на ДНК геномов, сложных клеточных структур и современных биохимических процессов жизни. Рождение синтеза белков было настолько важным для эволюции жизни, что Энтони Пул (A. Poole) и его коллеги из Стокгольмского университета (Швеция) назвали существо, у которого появился этот механизм, прорывным организмом, или Riborgis eigensis. Но возникновение генетически закодированного синтеза белков должно было «случайно» — или совершенно неожиданно — изменить химическую эволюцию на базе РНК. В самом деле, невозможно предвидеть какие-либо эволюционные «изобретения» до момента их неожиданного появления. Как мы знаем, для синтеза белков нужен сложный аппарат, состоящий из каталитических рибосомных комплексов (чтобы считывать генетический код и формировать пептидные связи), тРНК и аминёкислот, а еще требуются катализаторы, связывающие аминокислоты с тРНК (см. главу 28). Важнейшие компоненты этого механизма формируются молекулами РНК. Поскольку обе центральные каталитические функции, вовлеченные в процесс трансляции (аминоацилирование тРНК и формирование пептидных связей), могут управляться РНК-ферментами, то возможно, что исходная форма этого аппарата была создана РНК-миром. Но такие сложные механизмы не возникают вдруг, случайно; они должны развиваться, постепенно эволюционировать из некоторых ранее существовавших функций. Раз синтеза белков до этого не существовало, значит, исходный аппарат трансляции должен был развиться из чего-то другого. Группа Дэйвида Пенни из университета Массей (Новая Зеландия) предположила, что исходной функцией этого молекулярного механизма была репликация РНК и что все компоненты более позднего трансляционного аппарата клетки (проторибосомы, прото-тРНК и аминокислоты) были уже вовлечены в эту функцию. Организуя ранние функции этих компонентов, можно облегчить переход от их исходного взаимодействия к развитию процесса трансляции.
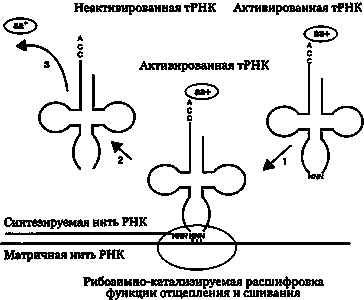
В соответствии с гипотезой группы Пенни механизм ранней репликации мог быть основан на активности рибосом-ферментов, или ранних рибосом, по расщеплению и сшиванию. В пользу этой гипотезы говорит то, что такая активность до сих пор очень распространена среди рибосом, а прямая полимеризация индивидуальных нуклеотидов — нет. Возможно, что предшественники рибосом опознавали целевые РНК-последовательности как триплеты нуклеотидов и копировали их в новую нить, сшивая вместе дополнительные триплеты, принесенные молекулами тРНК (рис. 30.6).
Рис. 30.6. Гипотетическая трансрепликация с помощью активированной тРНК. Исходная нить РНК показана внизу в горизонтальном виде. Рождающаяся реплицированная нить РНК показана внизу слева. Адаптировано из: Poole et al. 1998, The path from the RNA world. J. Mol. Evol. 46:1
Аминокислоты могли прикрепляться к тРНК, либо зарядив их энергией, либо обеспечив правильное складывание этих молекул. В процессе репликации аминокислоты приближаются друг к другу, и это позволяет формироваться пептидным связям между ними. Возможно, в течение некоторого времени ранний процесс репликации перекрывался ранним процессом трансляции. Эволюция генетически закодированных белков постепенно привела к возникновению белков, которые могли служить катализаторами репликации; эта функция была передана белковым ферментам, а рибосомы и тРНК превратились в чистый аппарат трансляции. Обе эти функции сохранились и укрепились в процессе естественного отбора.