Кортико-руброспинальная система двигательного контроля. От красного ядра, которое залегает в толще среднего мозга, берет начало руброспинальный путь. Моторная область коры, откуда берет начало пирамидный тракт, проецируется ипсилатерально на задние 2/3 красного ядра. Сенсомоторная кора (у кошки задняя часть сигмовидной извилины) проецируется ипсилатерально на переднюю область мелкоклеточной части красного ядра.
В ряду позвоночных животных структура красного ядра претерпевает существенные изменения (рис. 5.24). У рептилий и птиц оно однородно и содержит только крупные нейроны. У млекопитающих в дополнение к крупноклеточной (диаметр сомы 50–90 мкм) каудальной части ядра появляется ростральная часть, которая состоит преимущественно из мелких и средних нейронов (диаметр сомы 20–40 мкм). Соотношение этих двух разделов ядра варьирует, но у приматов увеличивается мелкоклеточная часть. Главный эфферентный путь красного ядра – руброспинальный тракт. После перекреста на уровне среднего мозга он спускается в мозговом стволе. В спинном мозге руброспинальный тракт идет в составе латеральных столбов (ростральнее латерального пирамидного тракта) и прослеживается до крестцовых сегментов спинного мозга. Основная часть афферентных проекций на мелкоклеточную часть красного ядра берет начало в зубчатом ядре мозжечка, а на мелкоклеточную часть – в промежуточных ядрах (круглом и пробковидном) мозжечка. Мелкоклеточное подразделение красного ядра образует восходящие проекции на вентролатеральное ядро таламуса, а нисходящие – на нижнюю оливу в продолговатом мозге. Увеличение размеров мелкоклеточной части и уменьшение крупноклеточной части красного ядра происходит на поздних стадиях эволюции и совпадает с появлением неомозжечковых структур и основной оливы. Регресс крупноклеточной части наиболее сильно выражен у человека и совпадает с неразвитостью руброспинального тракта.
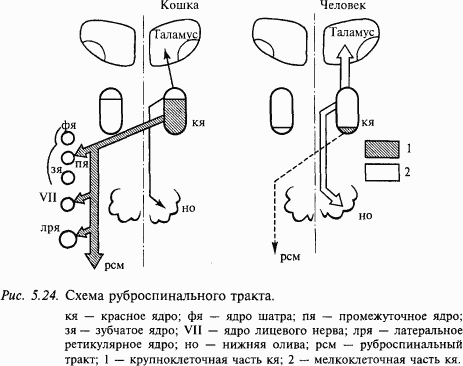
Рассечение ствола мозга у четвероногих (кошка, собака) на уровне ростральное переднего двухолмия не вызывает больших изменений в мышечном тонусе. Однако если разрез проходит на интраколликулярном уровне (изолируется красное ядро), это приводит у четвероногих животных к ригидности (так называемая децеребрационная ригидность). Она развивается по гамма-типу, т.е. обусловлена повышенной импульсацией в гамма-мотонейронах. В отличие от ригидности по альфа-типу эта ригидность пропадает после перерезки дорсальных корешков спинного мозга. Был сделан вывод, что крупноклеточная часть красного ядра осуществляет тормозное влияние на экстензоры постуральных мышц.
Опыты с электрической стимуляцией красного ядра показали, что преимущественно возбуждаются флексоры ипсилатеральной конечности. При стимуляции отдельных точек красного ядра прослеживается соматотопия.
В настоящее время есть основание предполагать, что обе системы двигательного контроля – кортико- и руброспинальные – организованы на уровне нейронного аппарата сегментарных механизмов спинного мозга в значительной степени сходно. Волокна обеих систем моносинаптически возбуждают в основном специализированные промежуточные нейроны латеральных участков серого вещества основания дорсального рога и нейроны промежуточного ядра и уже через эти переключения возбуждают мотонейроны.
Нейрофизиология ретикулоспинальных путей. Начало ретикулоспинальным путям дают нейроны на всем протяжении ретикулярной формации мозгового ствола. Эти пути спускаются в составе латеральных канатиков спинного мозга. Часть волокон в стволе мозга перекрещивается. Волокна ретикулоспинальной системы имеют различный диаметр и обеспечивают проведение возбуждения в широком диапазоне скоростей.
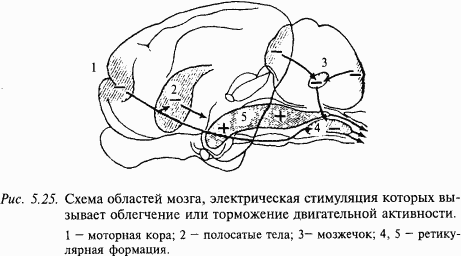
Облегчающие ретикулоспинальные влияния. Американские нейрофизиологи Р. Райнс и Г. Мэгун (1946) установили, что слабое электрическое раздражение ретикулярной формации мозгового ствола и более ростральных отделов мозга усиливает двигательные реакции на раздражение моторной области коры и заметно облегчает коленный рефлекс (рис. 5.25). Эти эффекты опосредованы ретикулоспинальными волокнами, преимущественно идущими в составе латерального канатика. Область, из которой вызываются двусторонние облегчающие влияния, включает значительную часть ретикулярной формации продолговатого мозга, покрышку моста и среднего мозга, центральное серое вещество, субталамус и гипоталамус, а также срединное и интраламинарные ядра таламуса.
Облегчающая ретикулоспинальная система играет важную роль в развитии децеребрационной ригидности и спастичности. На децеребрированных кошках (перерезка мозгового ствола на межколликулярном уровне) было показано, что стимуляция участков облегчающей системы ниже уровня перерезки усиливает и без того увеличенные в таких условиях проприоцептивные рефлексы. Этот эффект развивается билатерально. Влияние облегчающей системы мозгового ствола на экстензорный тонус было убедительно показано на кошках с хронической спастичностью, которая развивалась в результате удаления двигательной области коры больших полушарий. Спастическое состояние, устраняющееся во время эксперимента на фоне наркоза, не только восстанавливалось, но существенно усиливалось при стимуляции облегчающей системы мозгового ствола вплоть до развития судорог.
Тормозные влияния ретикулярной формации продолговатого мозга.
Значительный вклад в развитие представлений о нейрофизиологических механизмах контроля тонуса внесло открытие Г. Мэгуном и Р. Райнсом (1947) области вентральной ретикулярной формации продолговатого мозга, оказывающей тормозное влияние на разные виды двигательной активности: флексорный, коленный и другие рефлексы. Движения, вызванные раздражением моторной коры, также сильно тормозились. У децеребрированной кошки раздражение указанной области приводило к полной потере тонуса в ригидно разогнутых конечностях. Все тормозные эффекты проявлялись билатерально, но на ипсилатеральной стороне они имели меньший порог. Тормозные эффекты, вызываемые на спинальном уровне, были опосредованы ретикулоспинальными путями, проходившими в переднелатеральной области белого вещества спинного мозга. Активность тормозного механизма продолговатого мозга зависела от высших отделов мозга. При устранении этих влияний, например в результате высокой децеребрации и удаления мозжечка, ретикулоспинальный тормозный механизм переставал оказывать влияние на спинальную активность.
Электрическое раздражение тормозного центра мозгового ствола вызывает в мотонейронах сгибателей и разгибателей торможение. Нисходящие волокна, по которым осуществляется это влияние, проходят в вентральном квадранте спинного мозга в составе вентрального ретикулоспинального пути. Латентное время наблюдаемых эффектов указывает на их полисинаптичекую природу.
Другая группа тормозных эффектов опосредована дорсальной ретикулоспинальной системой, берущей начало примерно в тех же областях мозгового ствола, что и вентральный ретикулоспинальный тракт. После перерезки вентрального квадранта спинного мозга (для устранения влияний вентрального ретикулоспинального пути) стимуляция тормозного центра мозгового ствола уже не вызывает торможение в мотонейронах. По-видимому, непрерывный путь волокон в этой области вообще отсутствует, и тормозные эффекты осуществляются через проприоспинальные (собственные) волокна спинного мозга. Поэтому указанный путь обозначается как дорсальная ретикулоспинальная система.
Таким образом, наиболее древняя система надсегментарного моторного контроля ретикулярная формация мозгового ствола через ретикулоспинальные пути оказывает как облегчающие, так и тормозные влияния на весь нейронный аппарат спинного мозга. Наряду с быстропроводящими компонентами этих путей, обеспечивающими прямое действие ретикулярной формации на мотонейроны, волокна с меньшей скоростью проведения оказывают влияния на интернейронную сеть. Ретикулоспинальные пути иннервируют мотонейроны сгибательных и разгибательных мышц. Однако эти влияния оказываются преимущественно на проксимальные мышцы конечностей и мышцы туловища.