Наиболее ярким указанием на сверхтекучесть ядерного вещества является отличие моментов инерции сильно несферических ядер от твердотельных значений: теория сверхтекучести ядерного вещества удовлетворительно объясняет как абсолютные значения моментов инерции, так и их зависимость от параметра деформации Р. Теория предсказывает также резкое (скачкообразное) возрастание момента инерции в данной вращательной полосе при некотором критическом (достаточно большом) спине I . Это явление, аналогичное разрушению сверхпроводимости достаточно сильным магнитным полем, пока отчётливо не наблюдалось (в теоретическом предсказании критических значений I имеются неопределённости). Менее выразительно, но всё же заметно сказывается сверхтекучесть ядерного вещества на других свойствах ядра: на вероятностях электромагнитных переходов, на положениях оболочечных уровней и т. п. Однако в целом сверхтекучесть ядерного вещества выражена в реальных ядрах не так ярко, как, например, явление сверхпроводимости металлов или сверхтекучесть гелия при низких температурах. Причиной этого является ограниченность размера ядра, сравнимая с размером куперовской пары. Менее надёжны, чем в физике обычных конденсированных сред, и выводы теории сверхтекучести ядер. Главным препятствием теории и здесь является то обстоятельство, что взаимодействие между ядерными частицами не может считаться слабым (в отличие, например, от взаимодействия, приводящего к спариванию электронов в металле). Поэтому наряду с парными корреляциями следовало бы учитывать и корреляции большего числа частиц (например, четырёх). Вопрос о влиянии таких многочастичных корреляций на свойства ядра остаётся пока открытым.
Описанные ядерные модели являются основными, охватывающими свойства большинства ядер. Они, однако, не достаточны для описания всех наблюдаемых свойств основных и возбуждённых состояний ядер. Так, в частности, для объяснения спектра коллективных возбуждений сферических ядер привлекается модель поверхностных и квадрупольных колебаний жидкой капли, с которой отождествляется ядро (вибрационная модель). Для объяснения свойств некоторых ядер используются представления о кластерной (блочной) структуре Я. а., например предполагается, что ядро 6 Li значительную часть времени проводит в виде дейтрона и a-частицы, вращающихся относительно центра тяжести ядра. Все ядерные модели играют роль более или менее вероятных рабочих гипотез. Последовательное же объяснение наиболее важных свойств ядер на прочной основе общих физических принципов и данных о взаимодействии нуклонов остаётся пока одной из нерешенных фундаментальных проблем современной физики.
Лит.: Ландау Л. Д., Смородинский Я. А., Лекции по теории атомного ядра, М., 1955; Бете Г., Моррисон Ф., Элементарная теория ядра, пер. с англ., М., 1958; Давыдов А. С., Теория атомного ядра, М., 1958; Айзенбуд Л., Вигнер Е., Структура ядра, пер. с англ., М., 1959; Гепперт-Майер М., Йенсен И. Г. Д., Элементарная теория ядерных оболочек, пер. с англ., М., 1958; Мигдал А. Б., Теория конечных ферми-систем и свойства атомных ядер, М., 1965; Ситенко А. Г., Т артаковски и В. К., Лекции по теории ядра, М., 1972; Рейнуотер Дж., «Успехи физических наук», 1976, т. 120, в. 4, с. 529 (пер. с англ.): Бор О., там же, с. 545 (пер. с англ.); Моттельсон Б., там же, с. 563 (пер. с англ.).
И. С. Шапиро.
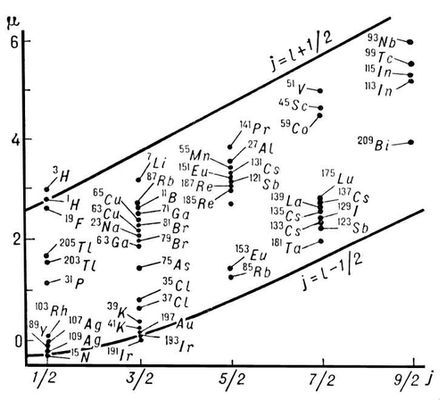
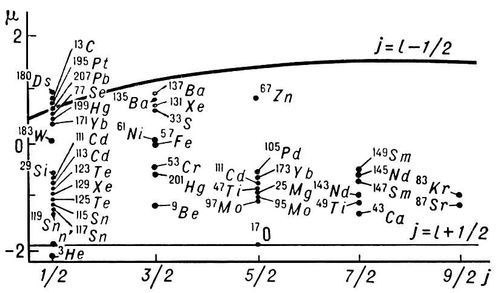
Рис. 2. Линии Шмидта для ядер с нечётным числом протонов (точки — экспериментальные данные).
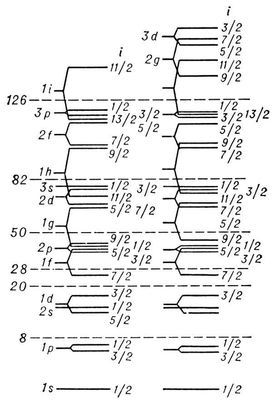
Рис. 3. Эмпирическая последовательность уровней протонов и нейтронов в модели ядерных оболочек. Справа от уровней указаны j, слева — спектроскопический символ (буква отвечает определённому значению l, число — номер уровня с данным l; s, р, d, f, g, h, i соответственно означают l = 0, 1, 2, 3, 4, 5, 6). Пунктиром отделены состояния, заполнение которых даёт магические числа.
Рис. 1. Линии Шмидта для ядер с нечётным числом нейтронов.
Ядро (биол.)
Ядро' клеточное, обязательная, наряду с цитоплазмой, составная часть клетки у простейших, многоклеточных животных и растений, содержащая хромосомы и продукты их деятельности. По наличию или отсутствию в клетках Я. все организмы делят на эукариот и прокариот . У последних нет оформленного Я. (отсутствует его оболочка), хотя дезоксирибонуклеиновая кислота (ДНК) имеется. В Я. хранится основная часть наследственной информации клетки; содержащиеся в хромосомах гены играют главную роль в передаче наследственных признаков в ряду клеток и организмов. Я. находится в постоянном и тесном взаимодействии с цитоплазмой; в нём синтезируются молекулы-посредники, переносящие генетическую информацию к центрам белкового синтеза в цитоплазме. Т. о., Я. управляет синтезами всех белков и через них — всеми физиологическими процессами в клетке. Поэтому получаемые экспериментально безъядерные клетки и фрагменты клеток всегда погибают; при пересадке Я. в такие клетки их жизнеспособность восстанавливается. Я. впервые наблюдал чешский учёный Я. Пуркине (1825) в яйцеклетке курицы; в растительных клетках Я. описал английский учёный Р. Броун (1831—33), в животных клетках — немецкий учёный Т. Шванн (1838—39).
Обычно Я. в клетке одно, находится близ её центра, имеет вид сферического или эллипсоидного пузырька (фигуры 1—3, 5, 6 ). Реже Я. бывает неправильной (фигура 4 ) или сложной формы (например, Я. лейкоцитов, макронуклеусы инфузорий). Нередки двух- и многоядерные клетки, обычно образующиеся путём деления Я. без деления цитоплазмы или путём слияния нескольких одноядерных клеток (т. н. симпласты, например поперечнополосатые мышечные волокна). Размеры Я. варьируют от ~ 1 мкм (у некоторых простейших) до ~ 1 мм (некоторые яйцеклетки).
Я. отделено от цитоплазмы ядерной оболочкой (ЯО), состоящей из 2 параллельных липопротеидных мембран толщиной 7—8 нм , между которыми находится узкое перинуклеарное пространство. ЯО пронизана порами диаметром 60—100 нм , на краях которых наружная мембрана ЯО переходит во внутреннюю. Частота пор различна в разных клетках: от единиц до 100—200 на 1 мкм2 поверхности Я. По краю поры располагается кольцо плотного материала — так называемый аннулус. В просвете поры часто имеется центральная гранула диаметром 15—20 нм , соединённая с аннулусом радиальными фибриллами. Вместе с порой эти структуры составляют поровый комплекс, который, по-видимому, регулирует прохождение макромолекул через ЯО (например, вход в Я. белковых молекул, выход из Я. рибонуклеопротеидных частиц и т. п.). Наружная мембрана ЯО местами переходит в мембраны эндоплазматической сети ; она обычно несёт белоксинтезирующие частицы — рибосомы . Внутренняя мембрана ЯО иногда образует впячивания в глубь Я. Содержимое Я. представлено ядерным соком (кариолимфой, кариоплазмой) и погруженными в него оформленными элементами — хроматином, ядрышками и др. Хроматин — это более или менее разрыхлённый в неделящемся Я. материал хромосом, комплекс ДНК с белками — так называемый дезоксирибо-нуклеопротеид (ДНП). Он выявляется с помощью цветной реакции Фёльгена на ДНК (фигуры 1 и 8 ). При делении Я. (см. Митоз ) весь хроматин конденсируется в хромосомы; по окончании митоза большая часть участков хромосом опять разрыхляется; эти участки (так называемый эухроматин) содержат в основном уникальные (неповторяющиеся) гены. Другие участки хромосом остаются плотными (так называемый гетерохроматин); в них располагаются главным образом повторяющиеся последовательности ДНК. В неделящемся Я. большая часть эухроматина представлена рыхлой сетью фибрилл ДНП толщиной 10 — 30 нм , гетерохроматин — плотными глыбками (хромоцентрами), в которых те же фибриллы плотно упакованы. Часть эухроматина также может переходить в компактное состояние; такой эухроматин считается неактивным в отношении синтеза РНК. Хромоцентры обычно граничат с ЯО или ядрышком. Есть данные о том, что фибриллы ДНП закреплены на внутренней мембране ЯО.