У растений, способных к «кооперативному» фотосинтезу, существует 2 типа Х., различающихся по строению и функциям. Одни из них, находящиеся в клетках мезофилла, мелкие с гранами, другие, более крупные, содержатся в клетках обкладки проводящих сосудистых пучков, граны в них лишь зачаточные или совсем отсутствуют. В Х. второго типа функционирует фотосистема 1, которая образует АТФ в ходе циклического фосфорилирования, а НАДФ×Н — за счёт реакции декарбоксилирования яблочной кислоты. Х. клеток обкладки фиксируют CO2 на рибулозодифосфате, т. е. с помощью цикла Калвина, а Х. клеток мезофилла — на фосфоенолпирувате (путь Хетча — Слэка); т. о. взаимодействие Х. обоих типов обеспечивает высокую эффективность фотосинтеза у растений. В строму Х., наряду с ферментами фиксации CO2 , включены нити ДНК, рибосомы, крахмальные зёрна, осмиофильные гранулы.
Наличие в Х. собственного генетического аппарата и специфической белоксинтезирующей системы обусловливает определённую, хотя и относительную, автономию Х. в клетке. При развитии и размножении растения в новых генерациях клеток Х. возникают только путём деления. Происхождение Х. связывают с симбиогенезом , полагая, что современные Х. — потомки сине-зелёных водорослей, вступившие в симбиоз с древними ядерными гетеротрофными клетками бесцветных водорослей или простейших.
Х. занимают 20—30% объёма растительной клетки. У водорослей, например хламидомонады, имеется один Х., в клетке высших растений содержится от 10 до 70 Х. Развиваются Х. из т. н. инициальных частиц, или пропластид, — небольших пузырьков, отделяющихся от ядра. В конце вегетации растения Х. в результате разрушения хлорофилла утрачивают зелёную окраску и превращаются в хромопласты . См. также Фотосинтез .
Лит.: Хлоропласты и митохондрии. Вопросы мембранной биологии, Сб., М., 1969; Лёви А., Сикевиц Ф., Структура и функция клетки, пер. с англ., М., 1971; Хит О., Фотосинтез, пер. с англ., М., 1972; Баславская С. С., Фотосинтез, М., 1974; Насыров Ю. С., Фотосинтез и генетика хлоропластов, М., 1975; Structure and function orchloroplasts, ed. М. Gibbs, B., 1971.
Р. М. Бекина.
Микрофотография хлоропласта.
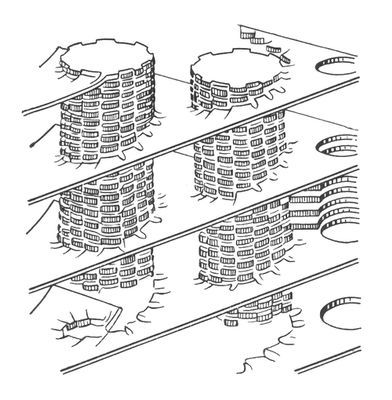
Модель пластинчатой (ламелярной) системы хлоропластов. Столбики — граны, образованные тилакоидами.
Хлоропрен
Хлоропре'н, 2-хлорбутадиен-1,3, CH2 =CH—CCl=CH2 , бесцветная жидкость с резким запахом; tkип 59,4 °С, плотность 0,9585 г/см3 (20 °С). Нерастворим в воде, смешивается с большинством органических растворителей. Х. присоединяет по двойным связям (обычно в положение 1,4) галогены, галогеноводороды, вступает в реакции диенового синтеза , чрезвычайно легко полимеризуется (поэтому его стабилизируют добавками пирогаллола или пирокатехина ). Получают Х. гидрохлорированием винилацетилена при 0—20 °С в присутствии хлорида меди Cu2 Cl2 и хлорида аммония; применяется для производства хлоропреновых каучуков. Токсичен; предельно допустимая концентрация в воздухе 0,002 мг/л.
Хлоропреновые каучуки
Хлоропре'новые каучу'ки', синтетические каучуки, полимеры хлоропрена общей формулы [—CH2 —CCl=CH—CH2 —] n ; продукты светло-жёлтого цвета. Плотность Х. к. 1,20—1,24 г/см3 , молекулярная масса (100—200)×103 , температура стеклования —40° С, удельное объёмное электрическое сопротивление 4,4×106 ом (м, электрическая прочность 23 Мв/м, диэлектрическая проницаемость 6,4—6,7. Для Х. к. характерен комплекс специфических свойств, обусловленных присутствием в их макромолекулах атомов хлора: масло-, бензо-, озоно- и теплостойкость, негорючесть, а также способность к вулканизации окислами металлов (в промышленности для этой цели применяют смеси ZnO и MgO). Х. к. кристаллизуются при растяжении, благодаря чему ненаполненные резины на их основе имеют высокую прочность. При наполнении Х, к. этот показатель резин в некоторых случаях снижается (см. Резина ), однако др. их ценные свойства, например сопротивление раздиру, бензостойкость, как правило, улучшаются.
Промышленный метод синтеза Х. к. — полимеризация в водной эмульсии. Основные области их применения — производство резино-технических. изделий, главным образом конвейерных лент, ремней, рукавов. Из Х. к. изготовляют также оболочки проводов и кабелей, защитные покрытия. Важное промышленное значение имеют клеи из Х. к. и хлоропреновые латексы . Мировое производство Х. к. ~ 400 тыс. т в год. Наиболее распространённые торговое название — наирит (СССР), неопрен (США).
Лит.: Энциклопедия полимеров, т. 3, М., 1977; см, также лит. при ст. Каучуки синтетические .
Хлорофилл
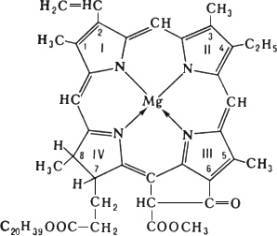
Хлорофи'лл (от греч. chlorós — зелёный и phýllon — лист), зелёный пигмент растений, с помощью которого они улавливают энергию солнечного света и осуществляют фотосинтез . Локализован в особых клеточных структурах — хлоропластах или хроматофорах и связан с белками и липидами мембран. Основу структуры молекулы Х, составляет магниевый комплекс порфиринового цикла; в IV пиррольном кольце к остатку пропионовой кислоты присоединён высокомолекулярный спирт фитол , который придаёт Х. способность встраиваться в липидный слой мембран хлоропластов.
Высшие растения и зелёные водоросли содержат Х. а и в , бурые и диатомовые водоросли — а и с , красные водоросли — Х. а и d. В фотосинтезирующих бактериях присутствуют близкие аналоги Х. — бактериохлорофиллы . По своему строению Х. близок к др. природным комплексам порфиринов (с железом) — дыхательным пигментам — цитохромам , красящему веществу крови — гему , а также простетическим группам некоторых ферментов — пероксидазы, каталазы.
Название «Х.» было дано французскими химиками П. Пельтье и Ж. Каванту зелёному спиртовому раствору смеси растительных пигментов в 1817. Впервые Х. а и в разделил в начале 20 в. рус. учёный М. С. Цвет с помощью разработанного им хроматографического метода. Химическую структуру Х. выяснили немецкие учёные Р. Вильштеттер , А. Штоль (1913), Х. Фишер (1930—40). Полный синтез Х. осуществил американский химик Р. Вудворд . Роль Х. в фотосинтезе доказана классическими работами К. А. Тимирязева . Пути биосинтеза Х. выяснены в трудах американских учёных Д. Шемина, С. Граника и др.; большой вклад в изучение Х. внесли советские учёные Т. Н. Годнев и А. А. Шлык.
Основной путь биосинтеза Х. определяется конденсацией двух молекул d-аминолевулиновой кислоты с образованием порфобилиногена — производного пиррола, который в результате ряда ферментативных превращений даёт соединение, содержащее порфириновое ядро — протопорфирин IX. Из протопорфирина образуется непосредственный предшественник Х. — протохлорофиллид, уже содержащий атом магния. Путём последующих реакций восстановления и присоединения фитола из этого предшественника образуется Х. Стадия восстановления протохлорофиллида осуществляется у высших растений на свету, у низших растений — в темноте.
В хлоропластах и хроматофорах большая часть Х. (содержание его обычно составляет 0,5—1,5% на сухую массу) находится в виде светособирающей «антенны» и меньшая часть — в реакционных центрах, непосредственно участвующих в работе цепи фотосинтетического переноса электрона. Поглощая квант света, молекула Х. переходит в возбуждённое состояние (длительность жизни синглетного возбуждённого состояния около 10-9сек ), которое может переходить в долгоживущее триплетное возбуждённое состояние с длительностью жизни до 10-3сек. Возбуждённые светом молекулы Х. способны переносить электрон от молекулы-донора к молекуле-акцептору. Механизм этих реакций в модельных системах выяснен в работах советских учёных А. А. Красновского, В. Б. Евстигнеева и др. Способность возбуждённого Х. к переносу электрона обеспечивает функционирование реакционных центров фотосистем цепи фотосинтетического переноса электрона. Применение спектральной техники и низких температур показало, что в первичном фотоакте бактериохлорофилл, а возможно, и Х. активного центра отдают свой электрон молекуле-акцептору (убихинон, ферредоксин). Этот первичный фотопроцесс сопряжён с цепью энзиматических реакций, ведущих к образованию восстановленных пиридиннуклеотидов и аденозинтрифосфата, обеспечивающих работу углеродного цикла. Т. о., свет, поглощённый Х., преобразуется в потенциальную химическую энергию органических продуктов фотосинтеза и молекулярного кислорода. Свет, поглощаемый Х., вызывает в клетках также др. фотобиологические явления: индуцирует генерацию электрического потенциала на мембранах хлоропластов, влияет на движение одноклеточных организмов (фототаксис) и т.д.