Деление клеток. В основе способности К. к самовоспроизведению лежат уникальное свойство ДНК самокопироваться и строго равноценное деление репродуцированных хромосом в процессе митоза. В результате деления образуются две К., идентичные исходной по генетическим свойствам и с обновленным составом ядра и цитоплазмы. Процессы самовоспроизведения хромосом, их деления, образования двух ядер и деления цитоплазмы разделены во времени, составляя в совокупности митотический цикл К. В случае, если после деления К. начинает готовиться к следующему делению, митотический цикл совпадает с жизненным циклом К. Однако во многих случаях после деления (а иногда перед ним) К. выходят из митотического цикла, дифференцируются и выполняют в организме ту или иную специальную функцию. Состав таких К. может обновляться за счёт делений малодифференцированных К. В некоторых тканях и дифференцированные К. способны повторно входить в митотический цикл. В нервной ткани дифференцированные К. не делятся; многие из них живут так же долго, как организм в целом, то есть у человека — несколько десятков лет. При этом ядра нервных К. не утрачивают способности к делению: будучи пересажены в цитоплазму раковых К., ядра нейронов синтезируют ДНК и делятся. Опыты с клетками-гибридами показывают влияние цитоплазмы на проявление ядерных функций. Неполноценная подготовка к делению предотвращает митоз или искажает его течение. Так, в некоторых случаях не происходит деления цитоплазмы и образуется двуядерная К. Многократное деление ядер в неделящейся К. приводит к появлению многоядерных К. или сложных надклеточных структур (симпластов), например в поперечнополосатых мышцах. Иногда репродукция К. ограничивается воспроизведением хромосом, и образуется полиплоидная К., имеющая удвоенный (сравнительно с исходной К.) набор хромосом. Полиплоидизация приводит к усилению синтетической активности, увеличению размеров и массы К.
Обновление клеток. Для длительной работы каждой К. необходимо восстановление изнашиваемых структур, как и ликвидация повреждений К., вызванных внешними воздействиями. Восстановительные процессы, характерные для всех К., связаны с изменениями проницаемости плазматической мембраны и сопровождаются усилением внутриклеточных синтезов, в первую очередь синтеза белка. Во многих тканях стимуляция восстановительных процессов приводит к репродукции генетического аппарата и делению К.; это свойственно, например покровам или кроветворной системе. Процессы внутриклеточного обновления в этих тканях выражены слабо, их К. живут сравнительно недолго (например, К. кишечного покрова млекопитающих — всего несколько суток). Максимальной выраженности внутриклеточные восстановительные процессы достигают в неделящихся или слабоделящихся клеточных популяциях, например в нервных К. Показателем совершенства процессов внутреннего обновления К. является длительность их жизни; для многих нервных К. она совпадает с продолжительностью жизни всего организма.
Мутации. Обычно процесс воспроизведения ДНК происходит без отклонений, и генетический код остаётся постоянным, что обеспечивает синтез одного и того же набора белков в огромном числе клеточных поколений. Однако в редких случаях может произойти мутация — частичное изменение структуры гена. Конечный её эффект — изменение свойств белков, кодируемых мутантными генами. Если при этом затрагиваются важные ферментные системы, свойства К., а иногда и всего организма существенно изменяются. Так, мутация одного из генов, контролирующих синтез гемоглобина, приводит к тяжелому заболеванию — анемии. Естественный отбор полезных мутаций — важный механизм эволюции.
Регуляция функций клеток. Основной механизм регуляции внутриклеточных процессов связан с различными влияниями на ферменты — высоко специфичные катализаторы биохимических реакций. Регуляция может осуществляться на генетическом уровне, когда определяется состав ферментов или количество того или иного фермента в К. В последнем случае регуляция может происходить и на уровне трансляции. Другой тип регуляции — воздействие на сам фермент, в результате чего может происходить как торможение, так и стимуляция его активности. Структурный уровень регуляции — влияние на сборку клеточных структур: мембран, рибосом и т.д. Конкретными регуляторами внутриклеточных процессов могут быть нервные влияния, гормоны, специальные вещества, вырабатываемые внутри К. либо окружающими К. (особенно белки), или же сами продукты реакций. В последнем случае воздействие осуществляется по принципу обратной связи, когда продукт реакции влияет на активность фермента — катализатора этой реакции. Регуляция может осуществляться через транспорт предшественников и ионов, влияния на матричный синтез (РНК, полисомы, ферменты синтеза), изменение формы регулируемого фермента.
Организация и регуляция функций К. на молекулярном уровне определяют такие свойства живых систем, как пространственная компактность и энергетическая экономичность. Важное свойство многоклеточных организмов — надёжность — во многом зависит от множественности (взаимозаменяемости) К. каждого функционального типа, а также от возможности их замены в результате размножения К. и обновления компонентов каждой К.
В медицине используются воздействия на К. для лечения и предупреждения заболеваний. Многие лекарственные вещества изменяют активность определенных К. Так, наркотики, транквилизаторы и болеутоляющие вещества снижают интенсивность деятельности нервных К., а стимуляторы её усиливают. Некоторые вещества стимулируют сокращение мышечных К. сосудов, другие — матки или сердца. Специальные воздействия на делящиеся К. осуществляются при использовании радиации или цитостатических веществ, блокирующих деление К. Иммунизация стимулирует деятельность лимфоидных К., вырабатывающих антитела к чужеродным белкам, предупреждая тем самым многие заболевания.
Лит.: Кольцов Н. К., Организация клетки, М. — Л., 1936; Вильсон Э., Клетка и её роль в развитии и наследственности, пер. с англ., т. 1—2, М. — Л., 1936—1940; Насонов Д. Н. и Александров В. Я., Реакция живого вещества на внешние воздействия, М. — Л., 1940; Кедровский Б. В., Цитология белковых синтезов в животной клетке, М., 1959; Мэзия Д., Митоз и физиология клеточного деления, пер. с англ., М., 1963; Руководство по цитологии, т. 1—2, М. — Л., 1965—66; Бродский В. Я., Трофика клетки, М., 1966; Живая клетка, [Сб. ст.], пер. с англ., М., 1966; Де Робертис Э., Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967; Васильев Ю. М. и Маленков А. Г., Клеточная поверхность и реакции клеток, Л., 1968; Алов И. А., Брауде А. И., Аспиз М. Е., Основы функциональной морфологии клетки, 2 изд., М., 1969; Лёви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971; Handbook of molecular cytology, ed. A. Lima-de-Faria, Amst., 1969.
В. Я. Бродский.
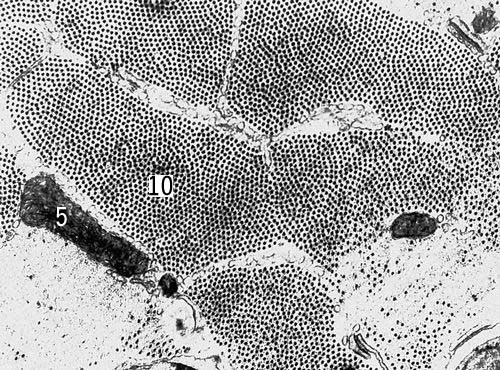
Поперечный срез мышечных клеток саранчи (увеличено в 54000 раз). Условные обозначения: 5 — митохондрии, 10 — миофибриллы.
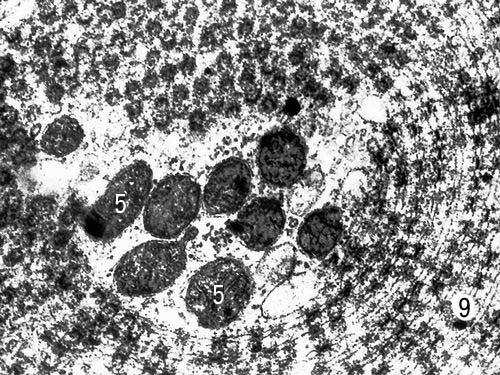
Специализированная форма мембран (пористые пластинки) в цитоплазме созревающей яйцеклетки севрюги (увеличено в 35000 раз). Условные обозначения: 5 — митохондрии, 9 — пористые пластинки.
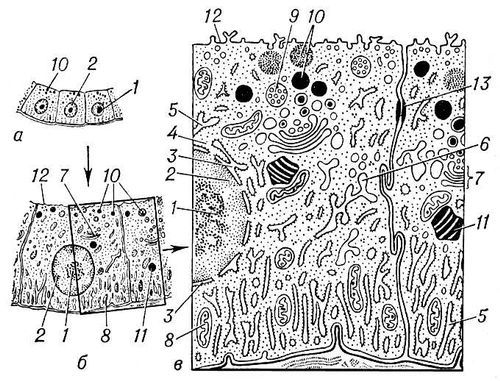
Рис. 1. Общий вид эпителиальной клетки животного при различном увеличении: а — в оптический микроскоп; б — при малом увеличении электронного микроскопа; в — при большом увеличении. Структуры ядра: 1 — ядрышко; 2 — хроматин (участки хромосом); 3 — ядерная оболочка. Структуры цитоплазмы: 4 — рибосомы; 5 — гранулярная (покрытая рибосомами) эндоплазматическая сеть; 6 — гладкоконтурная сеть; 7 — комплекс Гольджи; 8 — митохондрии; 9 — мультивезикулярные (многопузырьковые) тела; 10 — секреторные гранулы; 11 — жировые включения; 12 — плазматическая мембрана; 13 — десмосома.