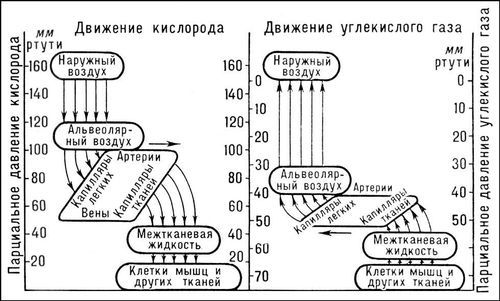
Лит.: Сеченов И. М., Избр. труды, М., 1935; Холден Дж. и Пристли Дж., Дыхание, пер. с англ., М.—Л., 1937; Маршак М. Е., Регуляция дыхания у человека, М., 1961; Физиология человека, М., 1966; Comroe J. Н., Physiology of respiration, Chi., 1966; Dejours P., Respiration, Oxf., 1966.
Л. Л. Шик.
Д. растений. Д. присуще всем органам, тканям и клеткам растения. Об интенсивности Д. можно судить, измеряя количество выделяемого тканью CO2 либо поглощаемого ею O2. Более интенсивно дышат молодые, быстро растущие органы и ткани растений. Наиболее активно Д. репродуктивных органов, затем листьев; слабее Д. стеблей и корней. Теневыносливые растения дышат слабее светолюбивых. Для высокогорных растений, адаптированных к пониженному парциальному давлению O2, характерна повышенная интенсивность Д. Очень активно Д. плесневых грибов, бактерий. Д. усиливается с повышением температуры (на каждые 10°С — примерно в 2—3 раза), прекращаясь при 45—50°С. В тканях зимующих органов растений (почки лиственных деревьев, иглы хвойных) Д. продолжается (с резко сниженной интенсивностью) и при значительных морозах. Д. стимулируют механические и химические раздражения (поранения, некоторые яды, наркотики и т.п.). Закономерно изменяется Д. в ходе развития растения и его органов. Сухие (покоящиеся) семена дышат очень слабо; при набухании и последующем прорастании семян Д. усиливается в сотни и тысячи раз. С окончанием периода активного роста растений Д. их тканей ослабевает, что связано с процессом старения протоплазмы. При созревании семян, плодов интенсивность Д. уменьшается.
Согласно теории советского биохимика А. Н. Баха, процесс Д., т. е. окисление углеводов, жиров, белков, осуществляется при помощи окислительной системы клетки в два этапа: 1) активирование O2 воздуха путём его присоединения к содержащимся в живой клетке ненасыщенным, способным самопроизвольно окисляться соединениям (оксигеназам) с образованием перекисей; 2) активирование последних с освобождением атомарного кислорода, способного окислять трудно окисляемые органические вещества. По теории дегидрирования русского ботаника В. И. Палладина, важнейшее звено Д. — активация водорода субстрата, осуществляемая дегидрогеназами. Обязательный участник сложной цепи процессов Д. — вода, водород которой вместе с водородом субстрата используется для восстановления самоокисляющихся соединений — так называемых дыхательных пигментов. CO2, выделяющийся при Д., образуется без участия кислорода воздуха, т. е. анаэробно. Кислород воздуха идёт на окисление дыхательных хромогенов, превращающихся при этом в дыхательные пигменты. Дальнейшее развитие теория Д. получила в исследованиях советского ботаника С. П. Костычева, согласно которым первые этапы аэробного Д. аналогичны процессам, свойственным анаэробам. Превращения образующегося при этом промежуточного продукта могут идти с участием кислорода, что свойственно аэробам. У анаэробов же эти превращения идут без участия молекулярного кислорода. По современным представлениям, процесс окисления, который составляет химическую основу Д., заключается в потере веществом электрона. Способность присоединять или отдавать электроны зависит от величины окислительного потенциала соединения. Кислород обладает самым высоким окислительным потенциалом и, следовательно, максимальной способностью присоединять электроны. Однако потенциал кислорода сильно отличается от потенциала дыхательного субстрата. Поэтому роль промежуточных переносчиков электронов от дыхательного субстрата к кислороду выполняют специфические соединения. Попеременно окисляясь и восстанавливаясь, они образуют систему переноса электронов. Присоединив к себе электрон от менее окисленного компонента, такой переносчик восстанавливается и, отдавая его следующему компоненту с более высоким потенциалом, окисляется. Так электрон передаётся от одного звена дыхательной цепи к другому и, в конце концов, кислороду. Таков заключительный этап Д.
Все эти процессы (активация кислорода, водорода, перенос электрона по цепи на кислород) осуществляются главным образом в митохондриях благодаря разветвлённой системе окислительно-восстановительных ферментов (см. Цитохромы). По пути следования к кислороду электроны, мобилизуемые первоначально от молекулы органического вещества, постепенно отдают заключённую в них энергию, которую клетка запасает в форме химических соединений, главным образом АТФ.
Благодаря совершенным механизмам запасания и использования энергии процессы энергообмена в клетке идут с очень высоким кпд, пока недостижимым в технике. Биологическая роль Д. не исчерпывается использованием энергии, заключённой в окисляемой органической молекуле. В ходе окислительных превращений органических веществ образуются активные промежуточные соединения — метаболиты, которые живая клетка использует для синтеза специфических составных частей своей протоплазмы, образования ферментов и др. Всем этим определяется центральное место, занимаемое Д. в комплексе процессов обмена веществ живой клетки. В Д. скрещиваются и увязываются процессы обмена белков, нуклеиновых кислот, углеводов, жиров и др. компонентов протоплазмы.
Лит.: Костычев С. П., Физиология растений, 3 изд., т. 1, М.—Л., 1937; Бах А. Н., Собр. трудов по химии и биохимии, М., 1950; Таусон В. О., Основные положения растительной биоэнергетики, М.—Л., 1950; Джеймс В. О., Дыхание растений, пер. с англ., М., 1956; Палладин В. И., Избр. труды, М., 1960; Михлин Д. М., Биохимия клеточного дыхания, М., 1960; Сент-Дьердьи А., Биоэнергетика, пер. с англ., М., 1960; Рубин Б. А., Ладыгина М. Е., Энзимология и биология дыхания растений, М., 1966; Рэкер Э., Биоэнергетические механизмы, пер. с англ., М., 1967; Рубин Б. А., Курс физиологии растений, 3 изд., М., 1971; Кретович В. Л., Основы биохимии растений, М., 1971.
Б. А. Рубин.
Рис. к ст. Дыхание.
Дыхания органы
Дыха'ния о'рганы, органы, служащие для газообмена между организмом и внешней средой. Д. о. имеются только у животных — аэробов, получающих свободный кислород или непосредственно из атмосферного воздуха (воздушное дыхание), или из воздуха, растворённого в воде (водное дыхание). У анаэробных организмов (одно- и многоклеточных) Д. о. отсутствуют. Многие группы низших аэробных организмов также не имеют Д. о. (простейшие, губки, кишечнополостные, многие черви). У животных с тонкими нежными покровами кислород может проникать в организм путём диффузии через всю поверхность тела — кожный тип дыхания, к которому часто присоединяется кишечное дыхание, когда кислород поступает в организм через стенку кишечного канала (например, у кишечнополостных). Среди позвоночных кожное дыхание играет большую роль у многих рыб и земноводных. Происхождение и строение Д. о. у разных организмов различны. Д. о. воздушного типа называются лёгкими и трахеями, Д. о. водного типа — жабрами.
Особый интерес представляют имеющиеся у рыб, живущих в иле (некоторых Clupeidae, Characinidae, Siluridae), дополнительные Д. о. — гроздевидные выросты, отходящие от жаберных дужек и служащие для воздушного дыхания. У лабиринтовых рыб на первой жаберной дужке развиваются костные пластинки, покрытые слизистой оболочкой, через которую поглощается O2. Это приспособление даёт возможность рыбе-ползуну (Anabas) долгое время находиться на суше.
У многих рыб из верхней части жаберных мешков могут вырастать воздушные пузыри, напоминающие настоящее лёгкое (у Amphipnous, Saccobranchus и др.). В процессе индивидуального развития у многих форм происходит смена различных Д. о. Так, среди насекомых у развивающихся в воде подёнок сначала имеет место диффузное кожное дыхание, заменяющееся впоследствии дыханием при помощи трахейных жабр, и, наконец, появляется свойственное взрослому насекомому трахейное воздушное дыхание. У личинок рыб до окончательного развития жабр дыхательную функцию выполняет вначале желточный мешок, снабжённый богатой сетью кровеносных сосудов, затем — сосуды непарных плавников, а иногда временные (личиночные) наружные жабры. У земноводных в личиночной стадии для дыхания служит сеть кровеносных сосудов желточного мешка, в дальнейшем обычно развиваются наружные жабры и часто возникают сосудистые сети в непарном (хвостовом) плавнике. Все зародыши амниот сначала дышат при помощи кровеносной сети желточного мешка. Впоследствии к такому дыханию присоединяется и вытесняет его так называемое аллантоидное дыхание, когда в стенках аллантоиса развивается густая сеть сосудов. Только после вылупления из яйца (пресмыкающиеся, птицы) или рождения (млекопитающие животные и человек) начинают функционировать их окончательные Д. о. — лёгкие (см. Дыхание).