При Д. дыхательная мускулатура преодолевает эластичное сопротивление, связанное с упругостью грудной клетки, тягой лёгких и поверхностным натяжением альвеол. Последнее, однако, значительно снижается под влиянием поверхностно активного вещества, вырабатываемого клетками альвеолярного эпителия; поэтому альвеолы при выдохе не спадаются, а при вдохе легко расширяются. Чем выше эластичное сопротивление, тем труднее растягиваются грудная клетка и лёгкие; при глубоком Д. работа дыхательной мускулатуры, затрачиваемая на его преодоление, резко возрастает. Неэластичное сопротивление Д. обусловлено главным образом трением воздуха при его движении по носовым ходам, гортани, трахее и бронхам. Оно зависит от скорости потока воздуха во время Д. и от его характера. При спокойном Д. поток близок к ламинарному (линейному) в прямых участках воздухоносных путей и к турбулентному (вихревому) в местах разветвления или сужения. С увеличением скорости потока (при форсированном Д.) турбулентность возрастает и для продвижения воздуха требуется более высокая разность давлений, а следовательно, и увеличение работы дыхательных мышц. Неравномерное распределение сопротивления движению воздуха по дыхательным путям приводит к тому, что поступление воздуха в разные группы лёгочных альвеол происходит неравномерно; такая разница в вентиляции особенно значительна при лёгочных заболеваниях.
Количество воздуха, вентилирующее лёгкие в 1 мин, называется минутным объёмом дыхания (МОД). МОД равен произведению дыхательного объёма на частоту Д. (число дыхательных движений в 1 мин, равное у человека примерно 15—18) и составляет у взрослого человека в покое 5—8 л/мин. Только часть МОД (около 70%) участвует в обмене газов между вдыхаемым и альвеолярным воздухом, эту часть называют объёмом альвеолярной вентиляции; остальная часть МОД используется для «промывания» так называемого мёртвого, или вредного, пространства дыхательных путей, в котором к началу выдоха сохраняется наружный воздух, заполнивший его в конце предшествовавшего вдоха (объём мёртвого пространства около 160 мл). Вентиляция альвеол обеспечивает постоянство состава альвеолярного воздуха. Парциальное давление O2(pO2) и CO2 (pCO2) в альвеолярном воздухе колеблется в очень узких пределах и составляет для О2 около 13 кн/м2 (100 мм рт. cт.) и для СО2 около 5,4 кн/м2 (40 мм рт. ст.).
Обмен газов между альвеолярным воздухом и венозной кровью, поступающей в капилляры лёгких, осуществляется через альвеоло-капиллярную мембрану, общая поверхность которой очень велика (у человека около 90 м2). Диффузия O2 в кровь обеспечивается разностью парциальных давлений O2 в альвеолярном воздухе и в венозной крови (8—9 кн/м2, или 60—70 мм рт. ст.). CO2, приносимый кровью из тканей в связанной форме (бикарбонаты, соли угольной кислоты и карбгемоглобин), освобождается в капиллярах лёгких при участии фермента карбоангидразы и диффундирует из крови в альвеолы; разность pCO2 между венозной кровью и альвеолярным воздухом составляет около 7 мм рт. ст. Способность альвеолярной стенки пропускать O2 и CO2, так называемая диффузионная способность лёгких, очень велика: в покое она составляет в 1 мин примерно 30 мл O2 на 1 мм разности pO2 между альвеолярным воздухом и кровью; для CO2 эта величина во много раз больше. Поэтому парциальное давление газов в оттекающей из лёгких артериальной крови успевает приблизиться к их давлению в альвеолярном воздухе. Переход O2 в ткани и удаление из них CO2 также происходят путём диффузии, т.к. pO2 в тканевой жидкости 2,7—5,4 кн/м2 (20—40 мм рт. ст.), а в клетках ещё ниже, а pCO2 в клетках может достигать 60 мм рт. cт. (см. рис.).
Потребление O2 клетками и тканями и образование ими CO2, что составляет сущность тканевого, или клеточного, Д., — одна из основных форм диссимиляции, осуществляющейся у животных и растений в принципе одинаково. Высокое потребление O2 характерно для тканей почек, коры больших полушарий головного мозга, сердца. В результате окислительно-восстановительных реакций тканевого Д. освобождается энергия, расходуемая на все жизненные проявления. Процесс этот осуществляется в митохондриях и складывается из дегидрирования субстратов Д. — углеводов и продуктов их расщепления, жиров и жирных кислот, аминокислот и продуктов их дезаминирования. Субстраты Д. поглощают O2 и служат источником CO2 (отношение —
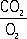
называется дыхательным коэффициентом). Энергия, освобождающаяся при окислении органических веществ, не используется тканями непосредственно, т.к. около 70% её расходуется на образование АТФ — одной из аденозинфосфорных кислот, последующее ферментативное расщепление которой обеспечивает энергетические потребности тканей, органов и организма в целом (см. Окисление биологическое, Окислительное фосфорилирование). Т. о., с биохимической точки зрения Д. — это превращение энергии углеводов и др. веществ в энергию макроэргических фосфатных связей.
Постоянство pO2 и pCO2 в альвеолярном воздухе, а стало быть, и в артериальной крови может поддерживаться лишь при условии, если альвеолярная вентиляция соответствует скорости потребления организмом O2 и образования CO2, т. е. уровню обмена веществ. Это условие обеспечивается благодаря совершенным механизмам регуляции Д. Управление частотой и глубиной Д. осуществляется рефлекторным путём. Так, повышение pCO2 и снижение pO2 в альвеолярном воздухе и в артериальной крови возбуждают хеморецепторы синокаротидной и кардиоаортальной зон, что приводит к возбуждению дыхательного центра и увеличению МОД. Согласно классическим представлениям, повышение pCO2 в артериальной крови, омывающей дыхательный центр, также возбуждает его и вызывает увеличение МОД. Т. о., регуляция Д. по отклонению pO2 и pCO2 в артериальной крови, осуществляемая по типу обратной связи, обеспечивает оптимальный МОД. Однако в ряде случаев, например при мышечной работе, МОД увеличивается до наступления в обмене веществ сдвигов, которые приводят к изменениям в газовом составе крови. Это усиление вентиляции обусловлено сигналами, поступающими в дыхательный центр от рецепторов двигательного аппарата, двигательной зоны коры больших полушарий мозга, а также условными рефлексами на различные сигналы, связанные с привычной работой и её обстановкой. Т. о., управление Д. осуществляется сложной самообучающейся системой не только по принципу регулирования по отклонению, но и по сигналам, предупреждающим о возможных отклонениях. Смена вдоха и выдоха обеспечивается системой взаимодополняющих механизмов. Во время вдоха в дыхательный центр по волокнам блуждающих нервов поступают импульсы от рецепторов растяжения, находящихся в лёгких. При достижении лёгкими определённого объёма эта импульсация тормозит клетки дыхательного центра, возбуждение которых вызывает вдох. При выключении нервных путей, обеспечивающих поступление импульсов в дыхательный центр, ритмичность Д. сохраняется благодаря автоматизму центра, однако характер ритма резко отличается от нормального. При нарушениях Д. и механизмов его регуляции возникают изменения газового состава крови (см. Гипоксия).
Методы исследования Д. разнообразны. В физиологии труда и спорта, клинической медицине широко применяют регистрацию глубины и частоты дыхательных движений, измерения газового состава выдыхаемого воздуха, артериальной крови, плеврального и альвеолярного давления. См. также Газообмен.