Градиенты концентраций всего двух факторов транскрипции могут породить более двух пространственных паттернов генной экспрессии. Активированный ген может кодировать белок, ответственный за онтогенетический процесс со строго определенным «адресом», либо другой фактор транскрипции, способный взаимодействовать с первыми двумя. Допустим, фактор C управляется схемой «много А и мало B» и распространяется путем диффузии (см. следующий рисунок). Тогда схема «много C и мало A» будет активна в узкой полосе сразу за правой границей области синтеза C – там, где А уже мало, а диффундирующего C еще достаточно, чтобы считать его концентрацию высокой.
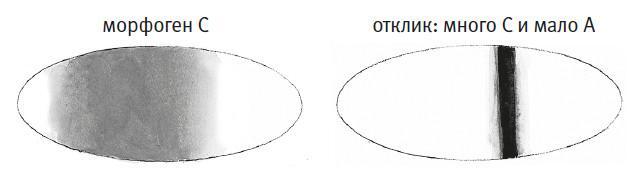
Узкая полоса иллюстрирует точность организации, обеспечиваемую небольшим числом морфогенов. Когда факторов транскрипции с характерными для них градиентами становится больше, спектр возможностей резко расширяется. Несложно представить, как можно задать специфические паттерны экспрессии генов, точно соответствующие назначению клеток, из которых формируются те или иные органы и ткани.
В теории это кажется убедительным, и организмы действительно применяют этот подход на практике. Процесс формирования паттернов мы уже несколько десятилетий приблизительно представляем и наблюдаем размытие концентраций факторов транскрипции и соответствующее профилирование экспрессии генов. В последние годы удается детальнее изучать эти процессы у все большего спектра организмов и с возрастающим числом вовлеченных генов. Лучше всего мы разбираемся в пространственной организации эмбрионов дрозофил, которых изначально исследовали уже знакомые нам Нюслайн-Фольхард и Вишаус. На ранних стадиях развития – задолго до появления ног, крыльев и даже головы – эмбрион мушки представляет собой тот самый удлиненный эллипс, что мы изображали чуть выше. У одного из полюсов внутри своей яйцеклетки (условно переднего, головного) мушка-мать оставляет наследство в виде мРНК, благодаря трансляции которой в зиготе создается стартовый градиент фактора транскрипции Bicoid, подобный градиенту нашего гипотетического морфогена B. Этот градиент задает передне-заднюю ось формирующегося тела. Bicoid связывается с промотором гена hunchback и активирует его экспрессию. Градиент концентрации белка Hunchback тоже ориентирован спереди назад. Далее широкими полосами экспрессируются еще полдюжины генов, комбинированные эффекты которых приводят к появлению более узких профилей экспрессии – например, семи полосок активности гена even-skipped[36], которые появляются всего через три часа после оплодотворения яйцеклетки.

Паттерны экспрессии even-skipped и других генов предопределяют разделение тела мушки на 14 сегментов. Из разных сегментов формируются разные структуры. Три сегмента, например, образуют грудной отдел, и на каждом из них при участии гена hedgehog вырастает по паре ног.
Особенно приятно, что наши биофизические знания позволяют измерять и предсказывать эти специфические паттерны полосок и сегментов, играющие важнейшую роль в определении плана тела любой зарождающейся дрозофилы. Что касается измерений, можно в живых эмбрионах исследовать транскрипцию интересующего гена, обеспечивая взаимодействие его РНК с фрагментами ДНК или РНК-связывающими белками, меченными флуоресцентным белком вроде GFP (см. главу 2): его легко локализовать по свечению. Если пришить ДНК флуоресцентного белка прямо к последовательности интересующего гена, можно по интенсивности свечения отслеживать количество его белкового продукта. В любом случае свечение служит точным, количественно измеримым показателем, позволяющим понять, что, где и когда происходит в онтогенезе9. Что касается прогнозирования, можно с помощью уравнений броуновского движения и функций генетического отклика рассчитать паттерны распределения белков и активности генов10. Эти расчетные данные пока не идеально отражают природные закономерности, но определяемые ими размеры и время появления полосок и зазоров довольно точно согласуются с реальной картиной мушиного эмбриогенеза.
Вообще плодовая мушка способна на гораздо более тонкую настройку в организации зародыша. В гипотетическом примере мы рассматривали лишь два уровня чтения гена – при низкой и при высокой концентрации морфогена. Но клетки могут обладать более утонченной чувствительностью и давать три разных ответа на три уровня морфогена (низкий, средний, высокий), а то и на четыре, пять и более. Казалось бы, неплохо различать больше уровней, ведь тогда организм станет высокоструктурированным за счет меньшего количества ингредиентов. Что же ограничивает точность эмбриона? Может ли плодовая мушка распознавать тысячу разных концентраций Bicoid в тысяче разных точек тела, формируя тысячу разных анатомических черт на базе единственного пространственного градиента?
Это тоже вопрос биофизики. Пределы точности такого структурирования задаются беспорядочностью диффузионного движения (см. главу 6) и хаотичностью молекулярного связывания, которую мы пока не рассматривали в деталях, но которая в основе своей схожа с броуновским движением. Несложно сказать, где в среднем окажется облако молекул в процессе диффузии, но насколько хорошо это среднее представляет все облако, зависит от количества движущихся молекул. Я совершенно уверен, что если подброшу миллион монеток, то получу примерно половину выпавших решек. Если же я подброшу шесть монеток, то ничуть не удивлюсь, увидев четыре решки и два орла. Подобным образом эмбрион может создавать обилие молекул морфогена, формируя плавный и четко очерченный градиент, который действительно позволяет вычленять множество разных уровней концентрации. Зародыш может производить и всего несколько молекул, тратя меньше сил и энергии, но это приводит к построению сильнее «зашумленных» градиентов, которые можно считывать лишь грубо – как области с высокой или низкой концентрацией. Судя по экспериментам, эмбрионы предпочитают скорее близкую ко второй стратегию: молекул морфогена не миллионы, и нечеткость градиентов существенна.
Пока неясно, как эта статистическая вариативность соотносится с точностью эмбрионального развития, но мы уже можем задаться вопросом, какие ограничения она накладывает на структурирование. В прекрасной статье 2013 года Уильям Бялек и его коллеги из Принстонского университета связали морфогенные измерения с теорией информации, чтобы выяснить, сколько битов закодировано в эмбрионе дрозофилы11. Если концентрация фактора транскрипции оценивается только как «высокая» или «низкая», ее можно закодировать в одном бите информации: мы отметили в первой главе, что один бит имеет два состояния. Если концентрация оценивается как «высокая», «средне-высокая», «средне-низкая» и «низкая», для кодирования четырех возможных состояний нам понадобится два бита. Неизвестно, сколько состояний активности каждого гена может различить регуляторная сеть мушки, и потому напрямую подсчитать число необходимых битов мы не можем. Бялек с коллегами, однако, поняли, что вариативность положений полосок и границ при сравнении разных эмбрионов отражает количество битов, используемых в структурировании. По сути, чем больше битов, тем выше точность и ниже вариативность, и наоборот. Анализ визуализаций эмбрионального развития дрозофилы, в частности паттернов экспрессии четырех генов, которые появляются вслед за паттерном bicoid и контролируют организацию тканей, показал, что на один ген приходится примерно два бита информации. Вместе четыре гена способны определять структуру с пространственной точностью около 1 %.
И снова мы сталкиваемся с поразительным свойством живого мира – созданием замечательных форм на основе скудного набора инструкций. Возможно, личинка не особо вписывается в ваши представления о «замечательном», но если мушки и не производят на вас впечатления, не забывайте, что эти податливые и удобные для изучения организмы позволили нам открыть и описать явления, которые оказались широко распространены. Вероятно, и вам сначала хватало нескольких битов.