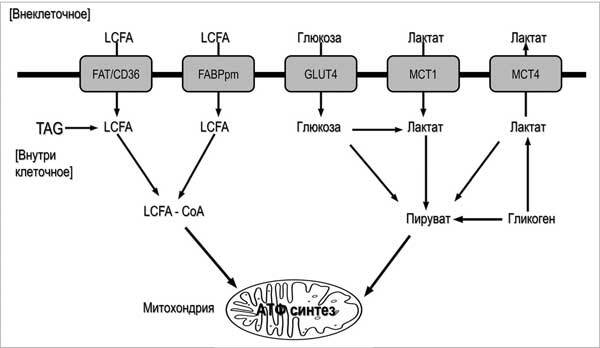
Рисунок 2.1. Упрощенная схема субстратного окислительного метаболизма (по данным D. Hoshinoetal., 2016, переработано). Обозначения: Длинноцепочечная жирная кислота, LCFA; триацилглицерин, ТАГ; транслоказа жирных кислот/CD36, FAT/CD36; белок, связывающий жирные кислоты, FABPpm; транспортер глюкозы 4, GLUT4; монокарбоксилатные белки-транспортеры, MCT.
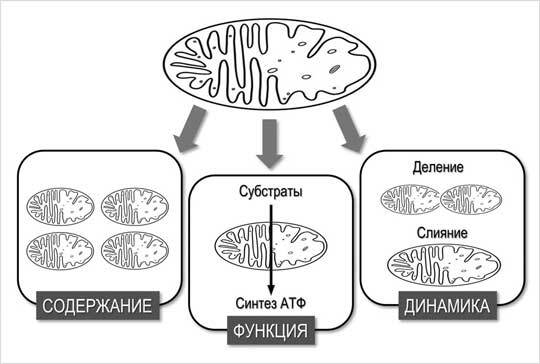
Рисунок 2.2. Митохондриальные адаптации: содержание, функция и динамика (по данным D. Hoshinoetal., 2016, переработано)
На рисунке 2.3 в общем виде представлен потенциальный механизм митохондриальной адаптации в скелетной мускулатуре. Высокоинтенсивные интервальные упражнения повышают концентрацию АДФ и АМФ, Са2+, АФК и лактата в скелетных мышцах. Внутриклеточные изменения окружающей среды, вызванные мышечными сокращениями, являются триггерами для активации сигнальной трансдукции, в том числе киназами, CaMK, AMPK и p38. Эти активации в киназах активируют белок PGC-1alpha и индуцируют его транслокацию в клеточные ядра. В ядре PGC-1alpha работает как котранскрипционный фактор, включая транскрипцию митохондриальных генов. Острых реакций недостаточно для увеличения содержания митохондриального белка, и для достижения долговременной митохондриальной адаптации цикл должен быть повторен.
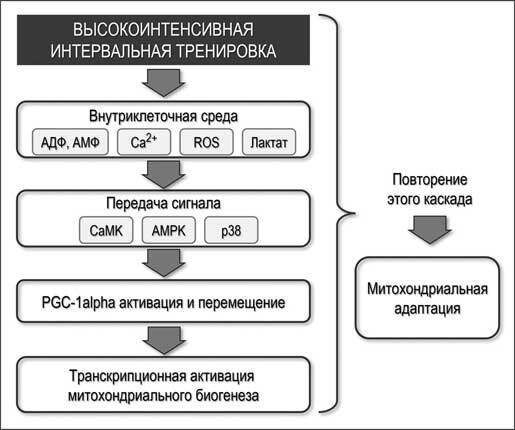
Рисунок 2.3. Общее представление потенциального механизма митохондриальной адаптации в скелетной мускулатуре (по данным D. Hoshino et al. (2016), переработано). Обозначения: Активные формы кислорода, АФК; Ca2+/кальмодулин-зависимая протеинкиназа, CaMK; АМФ-активируемая киназа, АМФК; митоген-активируемая протеинкиназа р38, р38
В аналитическом материале M. Atakan et al (2021) представлена краткая история высокоинтенсивных интервальных тренировок, основанная на новых результатах ряда исследований физической работоспособности и здоровья, начиная с начала 1920-х годов и до настоящего времени. Кроме того, проведен обзор механизмов, лежащих в основе физиологических адаптаций в ответ на высокоинтенсивные интервальные тренировки. Например, на рисунке 2.4 схематично показано, что упражнения с более высокой интенсивностью требуют большего «оборота» аденозинтрифосфата (А) и увеличивают высвобождение кальция из саркоплазматического ретикулума (B); окисление углеводов, особенно из мышечного гликогена, доминирует при более высокой интенсивности упражнений по сравнению с упражнениями с более низкой интенсивностью (C). Это приводит к большему накоплению метаболитов (аденозиндифосфат, аденозинмонофосфат, лактат, неорганический фосфат, креатин, кальций, ионы водорода, аденозинмонофосфатактивируемая протеинкиназа и кальций/кальмодулин-зависимая протеинкиназа II (D), вызывающие более высокие скорости экспрессии генов (E), что способствует более высоким скоростям синтеза митохондриального белка и большему содержанию митохондрий (F).
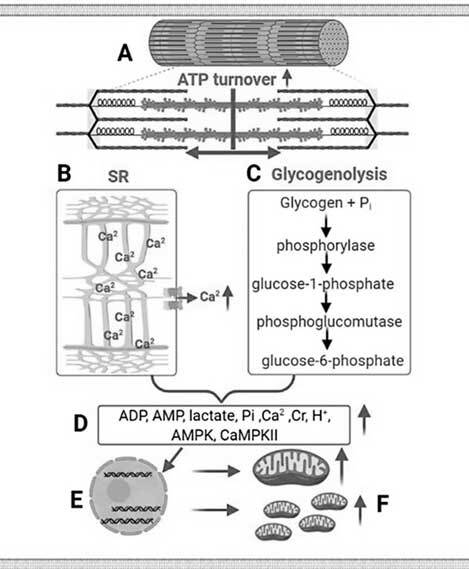
Рисунок 2.4 – Схема основных сигнальных путей, с помощью которых высокоинтенсивные упражнения вызывают большую митохондриальную адаптацию по сравнению с более низкой интенсивностью упражнений M. Atakan et al (2021). ADP-аденозиндифосфат (АДФ); AMP – аденозинмонофосфат (АМФ); AMPK, аденозин-монофосфатактивируемая протеинкиназа; ATP – аденозинтрифосфат (АТФ); Ca2, кальций; CaMPKII, кальций/кальмодулин-зависимая протеинкиназа II; Cr, креатин (Кр); H+, ион водорода; Pi, неорганический фосфат; SR; саркоплазматический ретикулум.
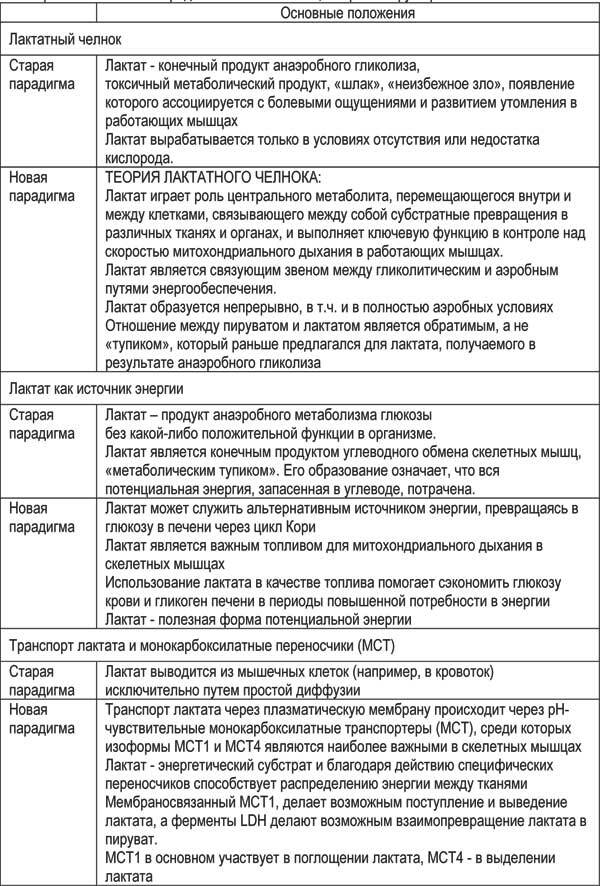
К важнейшим научным данным, получение которых не только существенно изменило многие широко распространенные представления, но и в значительной степени повлияло на методические подходы, в том числе и касающиеся программирования ВИИТ, безусловно, относятся результаты научных исследований последних десятилетий, направленных на изучение метаболической роли лактата.
Долгое время лактат считался токсичным побочным продуктом гликолиза, являющимся причиной усталости и отрицательно влияющим на результаты. Даже сегодня это мнение по-прежнему широко распространено. Однако, современные исследования определили лактату новое важное место и роль в метаболизме – это активный системный метаболит, перемещающийся внутри клеток, между клетками и между органами, способный не только использоваться для ресинтеза глюкозы (процесс глюконеогенеза), но и вовлекаться в энергетический обмен клеток, в том числе, окисляясь непосредственно в митохондриях (Brooks G. A., 2000; Ferguson, B. S., et al., 2018, Hashimoto T. et al., 2007.; О. В. Мещерякова и др., 2010, и др.). В работах G. Brooks et al. (1986, 2000, 2002 и др.) показано, что нормальные мышечные клетки производят лактат и в условиях покоя, и при умеренной нагрузке, а не только в условиях недостатка кислорода, т. е. значительная часть пирувата превращается в лактат, даже когда запас кислорода достаточен для поддержания аэробного метаболизма в митохондриях. (таблица 2.1).
Таблица 2.1 – Изменение представлений о лактате, его роли и функциях
Адаптация к тренировке состоит в повышении способности организма использовать лактат, окислять лактат как источник энергии, а не в повышении способности «терпеть» его наличие в гликолитических волокнах Поэтому важна возможность быстрого транспорта лактата, а, значит, совершенствования за счет тренировки механизмов этого транспорта Понимание внутриклеточных процессов помогает эффективнее строить тренировочный процесс и избегать факторов, нарушающих адаптацию.
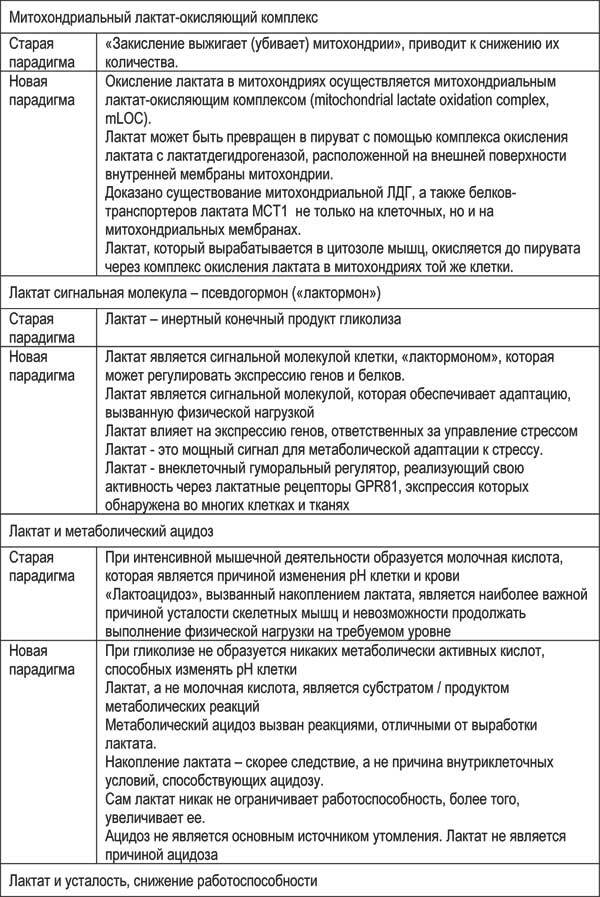
Смене концепции понимания роли лактата способствовал ряд работ, посвященных изучению внутри- и межклеточных систем переноса этого соединения (Gladden, L.B., 2004). Использование современных биохимических, иммунногистохимических, радиоизотопных и других методов позволило доказать существование механизма восстановления лактата до гликогена – внутриклеточного лактатного шаттла (intracellular lactate shuttle) (G. A. Brooks, 2018), с помощью которого лактат экзо-, а также эндогенного происхождения способен транспортироваться в митохондрии клеток скелетных мышц, сердца и нейронов и там подвергаться окислению. Отдельные детали этих механизмов в настоящее время еще являются предметом дискуссий, но общие принципы теперь ясны, и эти принципы имеют важное значение для оптимизации спортивной подготовки, прежде всего, в видах на выносливость.
Сегодня лактат уже не рассматривается как «побочный» или «вредный» продукт гликолиза. Результаты исследований показали, что окисление лактата является одним из самых важных источников энергии: в окислительных мышечных волокнах лактат является предпочтительным источником топлива (Brooks G. A., 1986). Было четко продемонстрировано, что межклеточный транспорт лактата осуществляется с помощью специальных белков-транспортеров – монокарбоксилатных переносчиков (monocarboxylate transporters – MCTs) (рисунок 2.5). Среди 14 идентифицированных изоформ MCT две – MCT1 и MCT4 – присутствуют в плазматических мембранах скелетных мышц, выявлена их связь с физической нагрузкой: физические упражнения увеличивают концентрацию в скелетных мышцах как MCT1, так и MCT4 (Kitaoka Y. et al, 2012).
В настоящее время доказано существование митохондриальной ЛДГ, а также белков-транспортеров лактата не только на клеточных, но и на митохондриальных мембранах (Hashimoto, Brooks, 2008; Hashimoto et all., 2008; Lemire et all., 2008).