А.Г. Пономаренко обращает в этой связи внимание на уже знакомые нам плавающие маты – гигантские скопления водных мхов и печеночников, обросших водорослями: клеточные стенки у этих растений не укреплены лигнином (в воде это не требуется), а главное – всё это битком набито животным белком в виде населяющих мат ракообразных и водных насекомых. Сходное с завроподами строение имеют зубы прочих ранних растительноядных групп (игуанодонов, стегозавров), так что не исключено, что все они вели приводный образ жизни, питаясь главным образом плавающими матами. Может статься, что наземными растительными кормами (более грубыми) динозавры стали питаться лишь во второй половине мела, когда среди них появились группы, имеющие зубные батареи – последовательные ряды плотно сидящих зубов (как у акулы), сменяющие друг друга по мере их стирания. Появление этих групп, приспособленных к питанию высокоабразивными кормами (гадрозавры, цератопсиды), было явно связано с распространением в это время покрытосеменных растений.
Итак, мезозой – время завроморфов: представители этой ветви доминируют и на суше (динозавры), и в морях (эвриапсиды), и в воздухе (птерозавры). А что же тероморфы? Именно в это время возникает самая продвинутая их группа, которая будет определять облик будущего, кайнозойского, сообщества позвоночных – млекопитающие (маммалии). Изучив детали этого процесса, Л.П. Татаринов (1976) выдвинул концепцию маммализации териодонтов – достаточно растянутое во времени появление у самых различных представителей таксона, рассматриваемого в качестве предкового, отдельных признаков, характерных для таксона-потомка. Было показано, что диагностические признаки, отличающие маммалий от рептилий (внутреннее ухо из трех косточек, мягкие губы, расширенные большие полушария мозга и т.д.) появляются в различных группах зверозубых ящеров параллельно и независимо, но лишь у одной группы – настоящих маммалий – возникает полный синдром из 8 таких признаков (рисунок 48). То есть, «стать млекопитающими» пытаются многие группы териодонтов, а кто именно достигнет результата (в нашем случае – потомки циногнат) – не столь уж важно.
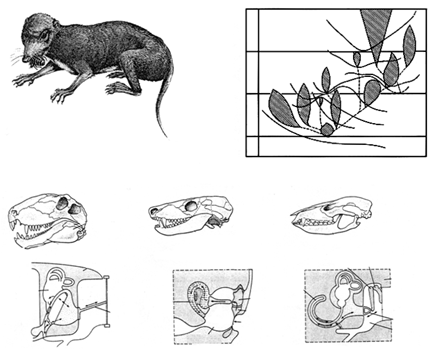
РИСУНОК 49. Происхождение млекопитающих: «маммализация териодонтов». (а) – Megazostrodon, древнейшее из известных млекопитающих из верхнего триаса; (б) – схема филогенеза по Татаринову (1976), демонстрирующая очередность приобретения маммальных признаков в различных филогенетических линиях тероморфов: I – звукопроводящий аппарат из трех слуховых косточек, II – вторичное челюстное сочленение между зубной и чешуйчатой костями, III – зачаточная барабанная перепонка в вырезке угловой кости. IV – мягкие, снабженые собственной мускулатурой губы, V – сенсорная зона на верхней губе (протовибриссы), VI – расширенные большие полушария головного мозга, VII – трехбугорчатые заклыковые зубы, VIII – верхние обонятельные раковины; (в) – эволюция челюстных структур и среднего уха (иллюстрирующая предыдущую схему).
Аналогичная тенденция параллельного развития ключевых признаков сохраняется и внутри маммалий. Так, среднее ухо «маммального типа», состоящее из трех слуховых косточек, входивших первоначально в состав нижней челюсти (у амфибий, рептилий и птиц есть единственная слуховая косточка, stapes), возникает у териевых млекопитающих (сумчатых и плацентарных) и прототериевых (однопроходных) млекопитающих двумя разными способами. Считается, что плацента у млекопитающих возникала минимум четырьмя различными способами (наш, «обезьяний» тип плаценты, когда зародыш буквально плавает в крови матери, – один из самых примитивных, по сравннению с тем, что существует, например, у грызунов); в последнем случае вполне правомочно говорить о плацентаризации териев (по аналогии с маммализацией териодонтов). По-видимому, таков вообще механизм происхождения всех крупных групп: вспомните тетраподизацию кистеперых рыб и «эукариотизацию».
В последнее время стало ясно, что происхождение птиц происходило по сходной схеме – ее можно назвать авиизацией архозавров. Птицы ведут свое происхождение от мелких ящеротазовых динозавров-манираптер, причем птичьи признаки тоже возникали неоднократно и независимо. Как полагает Е.Н. Курочкин (1996), полный синдром таких признаков возникал как минимум дважды: у хвостатых меловых энанциорнисов (они ведут свое происхождение от юрского археоптерикса, который, по сути, – еще просто оперенный динозаврик), и у настоящих, бесхвостых, птиц, чьими предками, судя по всему, были позднетриасовые тероподы протоависы (рисунок 47, д-е); беззубыми птицы становятся параллельно в обеих этих ветвях. Особенно же интересны такие «незавершенные» попытки авиизации, как небольшой – ростом с первоклассника – динозавр Avimimus, который имеет в скелете передней конечности пряжку (эта кость служит птицам для раскрытия маховых перьев) и череп с выставленными орбитами (это нужно, чтобы глаз не тонул в пере), или недавно описанный Sinopaleopteryx, покрытый чем-то вроде пухового пера (у археоптерикса перья только маховые). Можно предположить, что перьевой покров первоначально возникает как теплоизолятор для существ из малого размерного класса, не обладающих инерциальной гомойотермией, и лишь затем начинает использоваться для полета.
Вернемся, однако, к маммалиям. Часто пишут, что «мезозой был веком динозавров, а кайнозой – век пришедших им на смену млекопитающих»; это – типичный «научный предрассудок», из числа тех, о которых говорил С.В. Мейен (см. главу 9). На самом деле динозавры и млекопитающие появились на Земле одновременно – в конце триаса, и мирно сосуществовали потом на протяжении 120 миллионов лет. Более того, число известных на сегодня видов видов мезозойских млекопитающих просто-напросто превышает число видов динозавров; правда, все они были небольшими по размеру существами (по-настоящему маммалии вышли в крупный размерный класс только в эоцене), а потому не столь любимы популяризаторами науки и писателями-фантастами как гиганские рептилии, но это уже другой вопрос.