Однако против этой точки зрения выдвигалось серьезное – чисто палеонтологическое – возражение. Если процесс преобразования водорослей в высшие растения действительно происходил в прибрежных водах (где условия для попадания в палеонтологическую летопись наиболее благоприятны), то почему мы не видим никаких его промежуточных стадий? Тем более, что и сами харовые появляются в позднем силуре – одновременно с сосудистыми растениями, а особенности биологии этой группы не дают оснований оснований предполагать для нее длительный период «скрытого существования»... Поэтому появилась парадоксальная, на первый взгляд, гипотеза: а почему, собственно говоря, появление макроостатков высших растений в конце силура должно однозначно трактоваться как следы их выхода на сушу? Может быть, совсем наоборот – это следы переселения высших растений в воду? Во всяком случае, многие палеоботаники (С.В. Мейен, Г. Стеббинс, Г. Хилл) активно поддерживали гипотезу о происхождении высших растений не от водных макрофитов (типа харовых), а от наземных зеленых водорослей. Именно этим сухопутным (а потому не имеющим реальных шансов попасть в захоронения) «первичным высшим растениям» могли принадлежать загадочные споры с трехлучевой щелью, которые весьма многочисленны в раннем силуре и даже в позднем ордовике (начиная с карадокского века).
Впрочем, недавно выяснилось, что, судя по всему, правы сторонники обеих точек зрения – каждые по-своему. Дело в том, что некоторые из микроскопических наземных зеленых водорослей имеют тот же самый комплекс тонких цитологических признаков, что харовые и сосудистые (см. выше); эти микроводоросли теперь стали включать в состав Charophyta. Таким образом, возникает вполне логичная и непротиворечивая картина. Первоначально существовала – на суше – группа зеленых водорослей («микроскопические харовые»), от которой в силуре произошли две близкородственные группы: «настоящие» харовые, заселившие континентальные водоемы, и высшие растения, начавшие осваивать сушу, и лишь спустя некоторое время (в полном соответствии со схемой Мейена) появившиеся в прибрежных местообитаниях.
Из курса ботаники вам должно быть известно, что высшие растения (Embryophyta) делятся на сосудистые (Tracheophyta) и мохообразные (Bryophyta) – мхи и печеночники. Многие ботаники (например, Дж. Ричардсон, 1992) считают, что именно печеночники (если исходить из их современных жизненных стратегий) – главные претенденты на роль «первопроходцев суши»: они живут ныне на наземных водорослевых пленках, в мелководных эфемерных водоемах, в почве – совместно с синезелеными водорослями. Интересно, что азотфиксирующая сине-зеленая водоросль Nostoc способна жить внутри тканей некоторых печеночников и антоцеротовых, обеспечивая своих хозяев азотом; это наверняка было очень важно для первых обитателей примитивных почв, где этот элемент не мог не быть в жестком дефиците[34] . Упоминавшиеся выше споры из позднеордовикских и раннесилурийских отложений наиболее схожи именно со спорами печеночников (достоверные макроостатки этих растений появляются позже, в раннем девоне).
Однако в любом случае мохообразные (если даже они действительно появились еще в ордовике) облик континентальных ландшафтов вряд ли изменили. Первые же сосудистые растения – риниофиты[35] – появились в позднем силуре (лудловский век); вплоть до раннего девона (жединский век) они были представлены крайне однообразными остатками единственного рода Cooksonia, простейшего и архаичнейшего из сосудистых. А вот в отложениях следующего века девона (зигенского) мы находим уже множество разнообразных риниофитов (рисунок 30). С этого времени среди них обособляются две эволюционные линии. Одна из них пойдет от рода Zosterophylum к плауновидным (в их число входят и древовидные лепидодендроны – одни из основных углеобразователей в следующем, карбоновом, периоде). Вторая линия (в ее основание обычно помещают род Psilophyton) ведет к хвощеобразным, папоротникам и семенным – голосеменным и покрытосеменным (рисунок 30). Даже девонские риниофиты еще очень примитивны и, честно говоря, неясно – можно ли назвать их «высшими растениями» в строгом смысле: у них есть проводящий пучок (правда, сложенный не трахеидами, а особыми вытянутыми клетками со своеобразным рельефом стенок), но отсутствуют устьица. Такое сочетание признаков должно свидетельствовать о том, что растения эти никогда не сталкивались с дефицитом воды (можно сказать, что вся их поверхность являет собою одно большое открытое устьице), и, по всей видимости, являлись гелофитами (то есть росли «по колено в воде», вроде нынешнего камыша).
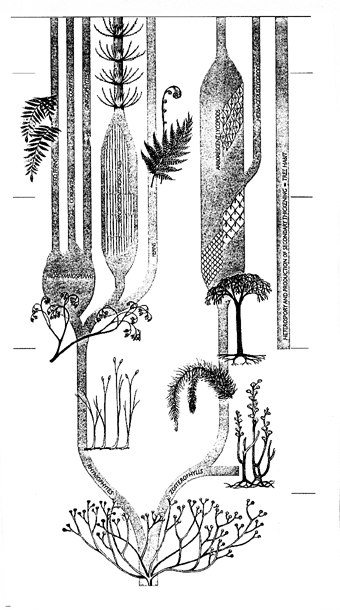
РИСУНОК 30. Родословное древо первых наземных растений.
Появление сосудистых растений с их жесткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы:
1. Фотосинтезирующие структуры стали располагаться в трехмерном пространстве, а не на плоскости (как это было до сих пор – в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества и, тем самым, суммарную продуктивность биосферы.
2. Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкоземом (по сравнению, например, с водорослевыми корками). Это уменьшило безвозвратные потери экосистемой неокисленного углерода (в виде органики) – совершенствование углеродного цикла.
3. Вертикальные стволы наземных растений должны быть достаточно жесткими (по сравнению с водными макрофитами). Для обеспечения этой жесткости возникла новая ткань – древесина, которая после гибели растения разлагается относительно медленно. Таким образом, углеродный цикл экосистемы обретает дополнительное резервное депо и, соответственно, стабилизируется.
4. Появление постоянно существующего запаса трудноразложимой органики (сконцентрированного в основном в почве) ведет к радикальной перестройке пищевых цепей. С этого времени большая часть вещества и энергии оборачивается через детритные, а не через пастбищные цепи (как это было в водных экосистемах).