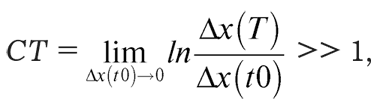9. Существует глубокая функциональная масштабная инвариантность, проявляющаяся в единой структуре принятия решений отдельными клетками при формировании тела нового организма, работой колонии насекомых и интегрированным поведением человека в обществе: это кибернетические процессы обучения и оптимизации параметров, реализуемые большим количеством субагентов, движимых инфотаксисом и стремлением к устойчивости гомеостаза.
10. Один из основных рычагов управления, используемых эгоистичными агентами, – динамика границ между их «индивидуальностью» и «окружающей средой». Передача сигналов между животными в экосистеме принципиально не отличается от передачи сигналов внутри мозга (или, в более широком охвате масштабов, внутри организма или внутри клетки) – все являются примерами информации, распространяющейся через сети локально-компетентных микроагентов, имеющих свои подвижные границы (Pais-Vieira et al., 2013; Kingsbury et al., 2019; Zhang and Yartsev, 2019).
Гипотеза безмасштабной когнитивности рассматривает опухоли многоклеточных как сжатие вычислительной границы клеточной биосистемы (потенциально обратимое): изолируя себя от физиологических сигналов окружающей ткани, когнитивная граница клетки сжимается до того небольшого размера, который свойственен одноклеточным. Раковые клетки не более эгоистичны, чем соматические клетки многоклеточного животного, но их индивидуальность теперь уменьшена до одной клетки, тогда как нормальные физиологические отношения в здоровых тканях связывают каждую клетку с общей целью организма. Майкл Левин считает, что принципы гипотезы в понимании опухолевого роста применимы не только к клеткам в органах, но и к скоплениям (роям) целых организмов, таких как пчелы и термиты (Seeley, 2009; Turner, 2011): в них наблюдается особая динамика сбоев в координации, имеющая существенные сходства, например между раком и колониями социальных насекомых. Также есть указания на параллели между динамикой рака и деградацией на уровне экосистем (Degregori and Eldredge, 2019). Обоснованно полагать, что аналогичный информационный подход применим и к другим патологиям как организмов, так и надорганизменных индивидуальностей, например роев, сообществ, организаций. То есть принципиально возможна система знаний, занимающаяся их «здоровьем» и «болезнями», которые вполне могут быть соотнесены с их инфекционными, метаболическими, опухолевыми аналогами на уровне организма.
От самогенерации цели к эгоистичности и индивидуальности элементов
Удивительно, насколько формулировки безмасштабной когнитивности Майкла Левина и автономной динамической информационной системы Д. С. Чернавского в применении к реальным биологическим системам перекликаются с концепцией «эгоистичного гена» Ричарда Докинза, который, как ни странно, в своих основных прорывных работах не опирается прямо на принципы статистической физики.
Важно понимать, что цель элемента (отдельного агента) отличается от целевой функции всей системы. Максимально общую целевую функцию можно предположить, на первый взгляд, как стремление к наиболее однородному (то есть термодинамически наиболее выгодному) состоянию, что в принципе может быть сообразно и цели отдельного элемента. Но в наиболее полной мере цели «выживших» («победивших») элементов и целевая функция всей системы могут совпадать лишь на последней стадии развития системы, когда почти все элементы обладают практически одинаковой информацией, и окончательное состояние системы полностью предсказуемо.
Однако время достижения этого состояния может стремиться к бесконечности, особенно когда есть непрерывный поставщик энергии в систему, и сама система является частью большой надсистемы. Более того, при условии постоянного поступления энергии в систему сама система может спонтанно самоусложняться и создавать иерархию (см., например, ниже теорию Джереми Ингланда). Также было показано, что в подобных моделях цели элементов могут изменяться в зависимости, скажем, от формирования новых иерархических над-уровней, а также может меняться (актуализироваться) тезаурус, то есть «своя информация» в зависимости от рецепции и генерации новой информации, когда новый случайный выбор делается в изменившемся контексте. Соответственно, имеющиеся ограничения в емкости «тары информации», особенно для сравнительно примитивных систем и их элементов, требуют избирательности в накоплении информации; минимально ценная информация в таких системах накапливаться не должна: в них просто может не хватить ни емкости «информационной тары», ни энергии, необходимой для ее поддержания.
Более развитые информационные системы, обеспечившие себе постоянный приток энергии и информации (в виде или рецепции, или генерации), могут позволить роскошь сохранять все менее ценную информацию, которая, с одной стороны, может оказаться чрезвычайно ценной в будущем, но, с другой стороны, служить источником противоречий с другими хранящимися информационными единицами, что, впрочем, может также служить и источником развития, что, в свою очередь, перекликается с концепцией цикла Дарвина-Эйгена (БОН: глава VI). В крайнем обобщении всю эволюцию материи можно представить, наверное, как непрерывное усовершенствование ее «информационной тары».
Как мы видим, задав определение «информационной системы», возможно прийти к выводу о способности самогенерации внутри ее первичной цели (ОП!). Согласно Д. С. Чернавскому, здесь важно взаимодействие всех трех важнейших аспектов: способностей к рецепции информации, генерации и сохранению. Так, для генерации информации необходимо сочетание как минимум двух условий: наличие хаоса (в математическом понимании, для случайности выбора) и механизма запоминания. Такое сочетание свойств есть по умолчанию далеко не у всех динамических систем, а только у тех, которые включают в себя, в терминологии Д. С. Чернавского, «перемешивающий слой». В самом общем определении «перемешивающий слой» – это область фазового пространства (то есть представления множества возможных состояний) мультистабильной (то есть потенциально способной находиться в нескольких устойчивых состояниях) динамической системы, обладающая следующими свойствами.
1. Все траектории системы (то есть последовательности смены состояний), исходящие из определенной области начальных условий, в момент времени t0 попадают в перемешивающий слой.
2. Все траектории в момент времени Т выходят из перемешивающего слоя и переходят в область мультистабильного динамического режима.
3. В области перемешивающего слоя имеет место стохастический (абсолютно случайный) режим, где произведение времени Т на величину С – аналога числа Ляпунова – много больше единицы:
где Δx(t0) и Δx(T) – расхождения траекторий в моменты входа и выхода из перемешивающего слоя, соответственно. Величина С переходит в соответствующее число Ляпунова при Т → ∞.
Числа Ляпунова (λ) – показатели, характеризующие устойчивость системы (математически – относящихся к ней странных аттракторов, см. ниже); если числа отрицательны, состояние системы устойчиво, если хотя бы одно из чисел положительно, то стационарное состояние системы неустойчиво.
Очень упрощая, можно сказать, что перемешивающий слой – та область фазового пространства, в котором неопределенность поведения объекта резко возрастает. В работах Г. Г. Малинецкого и соавторов (2000) области с подобными характеристиками называются областями со сверхбыстрой и малопредсказуемой динамикой или «джокерами» (jockers), а противоположные им по свойствам динамические области с медленной динамикой и хорошей предсказуемостью – «руслами» (channels), в которых неизбежно собираются множество изначально весьма различных траекторий. Последние области применительно к биологии можно сопоставить с креодами Конрада Уоддингтона (Conrad Waddington, 1964): «самоканализирующимися» траекториями развития организмов, которые определяются не только собственно генетической наследственностью, но и, например, ее эпигенетическим ландшафтом. При определенных условиях подобная канализация траекторий развития может выражаться в сходстве фенотипов при весьма различных генотипах, что может служить, например, причиной возникновения вавиловских гомологичных рядов у растений.