Соответствующие им степени свободы определяют собственные вибрации генома, которые взаимосвязаны в единую вибрационную матрицу. Геном с этих позиций является сложной многочастотной динамической машиной.
Собственные частоты f могут быть рассчитаны по известной формуле:
f = L/v,
где L – размер элемента структуры,
v – скорость распространений возмущения.
Это может быть скорость света, звука, движения зарядов или иного показателя. Также возможны варианты расчёта по энергиям межатомных и молекулярных связей, по формуле Де Бройля, по энергетическим затратам на разрушение структуры, по механическим характеристикам. В результате может быть определён спектр основных и производных от них частот для разных уровней структуры.
Помимо самостоятельной важности этих частот интересны и их взаимосвязи:
– между собой;
– со спектрами поглощения ДНК, ИК спектрами и т. д.;
– с характеристиками ЭМП, оказывающими значимое влияние на биообъекты. Если будет обнаружено совпадение, то оно будет свидетельствовать в пользу возможности резонанса с волновыми характеристиками Кода;
– с собственными частотами: воды, клеток, Земли, Луны, Вселенной (например реликтовое излучение).
Все эти предложения относятся к полям партнёров: РНК и белкам.
1.4.3. Измерения
Я предполагаю, что для измерения поля достаточно технологии секвенирования ДНК и высокоточных измерительных приборов. Кроме них в качестве датчиков могут использоваться биообъекты, различные физико-химические процессы, в том числе фазовые переходы.
Очень вероятно, что мы обнаружим иерархию полей с различными частотами, соответствующими различным уровням структуры ДНК, РНК, белков.
Роль генома определяют два различных процесса: акт оплодотворения с партнёром из иного организма и текущая авторепликация белков в процессе жизни. Динамика этих действий и соответствующие трансформации полей могут сильно различаться по интенсивности и длительности, но, вероятно, будут иметь сходную топологию по сути.
Рис. 2. Перенос действия образцов ДНК с помощью внешних полей на объекты: биосистемы, вещества, процессы
Изучать такой изменчивый и адаптивный объект, как поле генома, не внося при этом искажений в него, очень сложно. И всё же предложу несколько вариантов исследований:
– исследование динамики полей и излучений в фазе начала жизни (в момент оплодотворения) по аналогии с экспериментом сотрудников Массачусетского технологического института на примере морских звёзд (Patiria miniata) (подробности работы в п. 1.7);
– модуляция внешних полей (ЭМ, акустическое) и излучений образцами ДНК и перенос этого действия на различные вещества и процессы, в том числе биосистемы (рис. 2);
– отличие в полях образцов ДНК, извлечённых из организма, и непосредственно в «рабочем состоянии» внутри него;
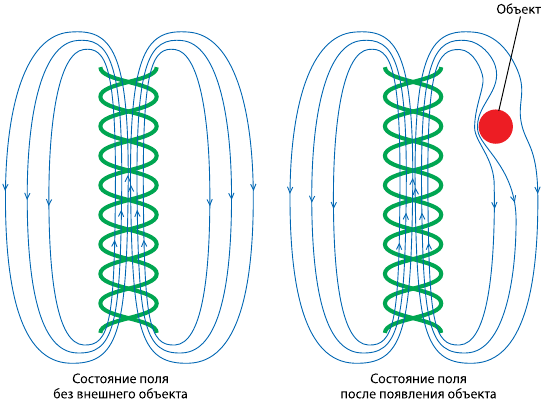
– изменения в геноме и его поле под влиянием внешнего искажения (рис. 3);
– изменение в поле образца Кода после изменения его структуры, например после удаления какого-либо звена;
– использование техники Кирлиан для массивных образцов ДНК;
– усиление, фокусировка, отклонение и другие манипуляции с полями Кода;
Рис. 3. Изменения в поле генома под влиянием внешнего объекта
– изменение направления и условий переноса генетической информации под влиянием внешнего поля;
– анизотропия полей образцов ДНК со структурой, выстроенной в одном направлении; возможно усиление эффекта внешними полями;
– направление вращения поля;
– поле как стоячая волна;
– поля ДНК, влияющие на квантовый уровень: отклонение частиц, дифракция и т. д.;
– поля белков, РНК и др.;
– поля как детектор движений и временной динамики генома;
– диагностика эволюции структуры и поля генома короткоживущих объектов методами атомной микроскопии или чем-то в этом роде в режиме реального времени;
– геном – высокоселективная система. Его действие и чувствительность будут проявляться в узких частотных интервалах. Поэтому, применяя соответствующие датчики и излучатели, его можно уверенно выделять в спектре сопутствующих полей. Например с помощью известных методов спектроскопии. Вероятно, удастся разработать метод пространственной диагностики, по типу МРТ. И если современная МРТ основана на фиксации изменений состояний атомов водорода, то, возможно, и для ДНК найдётся соответствующий элемент или молекула. Для этого приемлемы методы изучения магнитных материалов, веществ, содержащих магнитные кластеры, и т. д. В результате мы визуализируем геномное поле, его дефекты, неоднородности и т. д. Оно может быть методом диагностики отдельной клетки, органа и организма в целом. Можно предположить, что поле здорового организма будет существенно отличаться от повреждённого или больного. Интересно, обнаружим ли мы при этом голографическую структуру, о которой так много говорилось?
– исследование состояний внутренней воды, контактирующей с геномом.
Открытой для меня остаётся проблема прямого эксперимента, доказывающего, что часть генетической информации содержится в самой геометрической структуре. Можно попытаться деструктурировать ДНК до простой линейной последовательности, вытянутой в линию, растянув спиральную структуру с помощью поля, и оценить влияние на процесс репликации. Но я не представляю, возможно ли это вообще без нарушения естественной структуры цепочки. Результаты можно сопоставить с известным действием факторов, искажающих генетическую информацию или её передачу.
1.4.4. Использование эффекта формы
Под эффектом формы я понимаю возможность влияния на состояние и характеристики объекта некой вещественной геометрической формы, не являющейся его частью, внешней по отношению к нему. Не имею возможности и смысла представлять целиком всю свою отдельную работу, посвящённую описанию этого эффекта. Ниже изложены тезисы из неё.
Внешняя ограничивающая форма
Примеры: фигуры Хладни, процессы внутри пирамид, рост кристаллов, явления самоорганизации и т. д. Свежий пример из биологии – экспериментально установленный эффект перепрограммирования стволовых клеток в клетки другого типа путём изменения формы ограничивающей площадки, на которой они выращивались. При этом специальные молекулярные регуляторы не использовались (R. Venkatachalapathy et al. PNAS, 2018, v. 115, n. 21, p. 4741–4750).
Т.е. макроформа в ряде случаев может определять структуру на микроуровне. Наши представления позволяют сформулировать несколько вариантов объяснения этого эффекта:
– поля Кода в колониях клеток образуют единую полевую структуру и начинают чувствовать ограничивающие их формы. Это заставляет систему меняться в целом и в каждой её части-клетке. Совершенно невероятно, чтобы такое влияние могло быть распознано одной клеткой. Это синергетический эффект, приводящий к переходу на новый качественный уровень. Объединять клетки вместе может, вероятно, только дальнодействующее поле;
– вибрации и излучения, порождаемые клетками, отражаются от стенок формы, возвращаются назад и стимулируют собственные процессы. Это эффект автостимуляции;
– внешние вибрации, попадающие на ограничивающую форму, формируют особый энергетический ландшафт по типу фигур Хладни.
Возможно повторить этот эффект на больших масштабах, дабы избежать прецизионных измерений. Для этого можно использовать представителей обеих видов материи: бактерии, дрожжи, монокультуры мелких объектов, кристаллы, полимеры и т. д. Важна зависимость эффекта от масштаба. Для усиления форму можно изготавливать из материалов, оказывающих сильное воздействие на ЭМП, с частотами, близкими к собственным вибрациям клеток.