T-лимфоциты используют свои рецепторы на поверхности клетки для взаимодействия с белковыми фрагментами или пептидами вирусных антигенов, прикрепленными к MHC на поверхности инфицированных клеток. Эти белки MHC в действительности переносят вирусные пептиды на поверхность клеток. Таким образом, T-лимфоциты ищут чужеродные антигены (в данном случае вирусные антигены – пептиды, произведенные из вирусного антигена) на поверхности зараженных клеток, на которых данный вирус паразитирует. Цитотоксические T-клетки, опознав инфицированную клетку как «чужеродную» (содержащую вирус), активируются и немедленно уничтожают зараженную клетку и/или высвобождают хемокины и цитокины, предупреждающие об опасности и вооружающие другие клетки организма хозяина на борьбу с вирусом. Кроме того, некоторые из этих цитокинов способны непосредственно препятствовать репликации вирусов. Такими средствами распространение вируса ингибируется, и очаг инфекции ликвидируется.
Иммунный ответ антител и цитотоксических T-клеток зависит от количества лимфоцитов, которые производятся кроветворными стволовыми клетками во время процесса образования крови. Антитела и CTL представляют две составляющие специфических реакций иммунной системы на антигены; оба они играют важную роль в борьбе с инфекцией. На самом деле вся иммунная система обладает врожденной гибкостью, благодаря которой соответствующий вклад каждой из ее составляющих варьируется в зависимости от природы инфицирующего вируса. Антитела в основном воздействуют на вирусы в жидкостных средах тела, и, соответственно, наиболее эффективно ограничивают их распространение в крови или цереброспинальных жидкостях, которые омывают головной и спинной мозг. Посредством этого антитела уменьшают содержание вирусов и снижают их способность инфицировать клетки, а следовательно, и количество зараженных клеток. Однако уничтожение инфицированных вирусом клеток и их удаление из организма – дело главным образом CTL. Расправляясь с инфицированными клетками, T-лимфоциты ликвидируют производство вирусного потомства. По мере того, как число вирионов сокращается, работа антител становится проще.
Прежде чем продолжить рассказ о T-лимфоцитах, я сделаю отступление, чтобы описать антитела, которые являются большими белковыми молекулами. Антитела производятся В-лимфоцитами, названными так из-за их источника – костного мозга1,2. В-клетки – это малые лимфоциты, находящиеся в состоянии покоя, с ядром, которое практически заполняет всю клетку; цитоплазмы в них незначительное количество. Когда вирус или вирусный антиген встречается со специфическим лимфоцитом с заранее подготовленным рецептором для антитела, которое соответствует белковой структуре вируса, В-лимфоцит получает стимул к делению, и количество цитоплазмы, заполняющей объем клетки, увеличивается, то есть В-лимфоцит дифференцировался в плазматическую клетку. В увеличившейся в объеме цитоплазме начинают производиться и немедленно отправляться в бой антитела, сконструированные для воздействия на тот самый вирус, который дал толчок к их секреции. Один-единственный В-лимфоцит в состоянии выпустить 100 миллионов антивирусных антител в час.
Антитела захватывают и нейтрализуют вирусы, используя один из нескольких механизмов1,16:
1) антитела могут предотвратить заражение, обволакивая внешний шиповидный белок вируса, который закрепляется на рецепторе клетки и дает возможность вирусу проникнуть внутрь;
2) антитела могут собрать вирусы вместе, в комок, так что количество нетто инфекционных частиц сократится;
3) с помощью комплемента/подкрепления – группы белков в крови – антитела могут лизировать (разрушать) вирусы16; и
4) антитела могут воздействовать на вирусные антигены на внешней мембране инфицированной клетки, чтобы ограничить производство или транскрипцию вирусных молекул внутри клетки, а следовательно, сократить количество создаваемых вирусов17.
Каждая молекула антитела воздействует на конкретный антиген или определенную вирусную молекулу-мишень. С помощью генов, дающих указания синтезу разнообразных антител, организм-хозяин может синтезировать их миллиарды. Развитие и поддержание в рабочем состоянии антивирусных антител и T-клеток происходило для того, чтобы они действовали согласованно для сдерживания вирусных инфекций. Однако при определенных вирусных заболеваниях (корь, грипп, ВИЧ, и т. д.) основная роль ложится на T-клетки, тогда как при взаимодействии с семейством энтеровирусов (полиомиелит, вирусы Коксаки) главная роль оказывается у антител. Трансфер антител может ограничить развитие текущей инфекции, например, при ВИЧ18. Это происходит, вероятнее всего, следующим образом. Во время инфекций с высокой вирусной нагрузкой на организм-хозяин вирус истощает антивирусные T-клетки. Применение антивирусных антител снижает вирусную нагрузку, тем самым восстанавливая функционирование T-клеток (облегчение истощенного состояния). После этого функционирующие антивирусные T-клетки могут уничтожить клетки-фабрики, реплицирующие вирус.
Теперь, возвращаясь к T-клеткам, следует отметить, что CD4+ T-лимфоциты в некоторых особых случаях также действуют как цитотоксические клетки. И наоборот, хотя их основная роль – распознавать и уничтожать зараженные вирусом клетки, CD8+-клетки могут высвобождать растворимые молекулы, поэтому они обладают функцией хелперов/индукторов. В тканевой культуре одна клетка CD8+ T-лимфоцита может убить до 10 и даже больше инфицированных вирусом клеток, захватив зараженную клетку и растворив ее, а затем, перейдя к следующей инфицированной клетке, повторить процесс и т. д. (Рисунок 3.1.) Далее в организме живых подопытных животных один CD8+ T-лимфоцит, как показало наблюдение, единовременно связывал и разрушал до трех зараженных клеток-мишеней19. При этом очень важно то, что эти T-лимфоциты могут распознавать вирусные пептиды на инфицированных клетках прежде, чем закончится сборка вируса, и эффективно и быстро уничтожать эти клетки до образования вирионов.
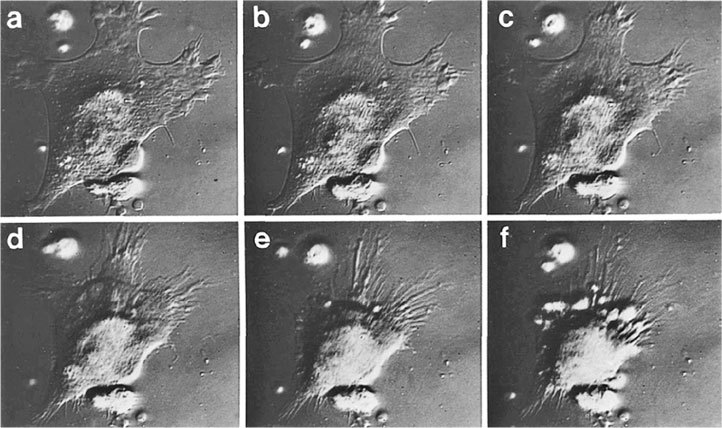
РИСУНОК 3.1. Стадии уничтожения инфицированной вирусом клетки Т-лимфоцитами (a – c): (а) стрелка указывает на Т-лимфоцит, прикрепившийся к зараженной клетке; (с) инфицированная вирусом клетка сжалась, на ней появились волдыри или вздутия, и она погибла. Все эти стадии проходят менее, чем за две минуты. Бар, 14 мк; (d – f) прикрепление Т-лимфоцита к клетке-мишени, которую лимфоциты уничтожить не могут. Продолжительность всего процесса от (d) до (f) – более 30 минут. Бар, 8 мк. Микрофотография Клауса Хана и Майкла Б. А. Олдстоуна.
Когда организм впервые сталкивается с инфицирующим его вирусом или подвергается воздействию вакцины, содержащей вирусные антигены, он генерирует специфические антитела, а также цитотоксические лимфоциты, реагирующие именно на этот вирус. (Рисунок 3.2.) Реакция CTL-лимфоцитов начинается в первый день заражения и увеличивается в 105–106 раз посредством удвоения примерно каждые 12 часов; пик роста достигается на седьмой-восьмой день после заражения.
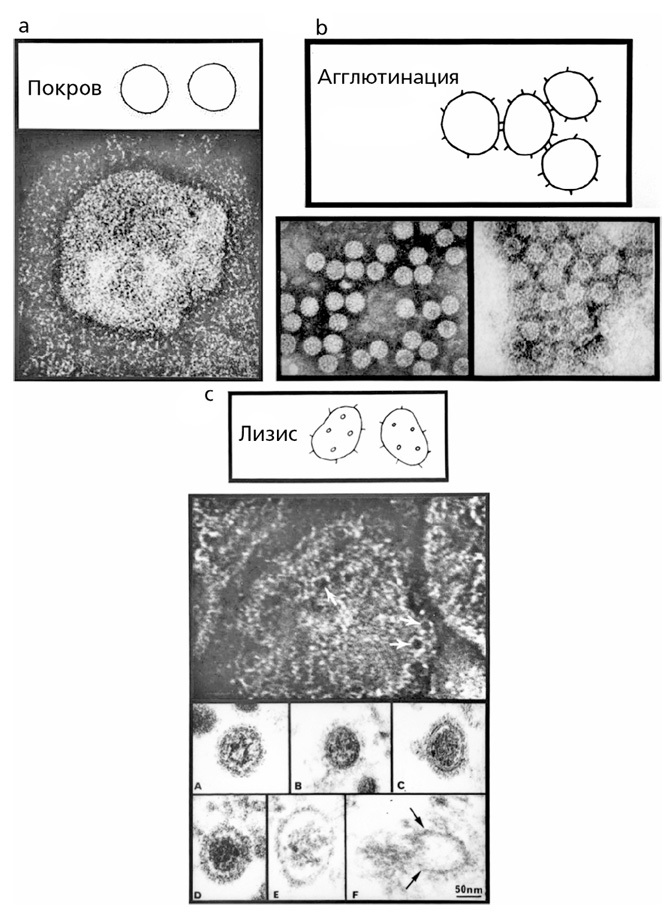
РИСУНОК 3.2. С помощью дополнительных белков (протеинов) в крови или без них антитела могут обволакивать вирусы: (а) антитело воздействует на коронавирус, собирая вирусные части вместе в один ком; (в) антитело при помощи комплементарных агентов немедленно разрушает вирус; (с, вверху) ретровирус, (с, внизу) аренавирус. В процессе разрушения ретровируса в нем образуются дыры (стрелки на рисунке), однако лизис (разрушение/растворение) аренавируса запускает цепочку событий, которые в конечном итоге приводят к выходу вирусных нуклеиновых кислот во внешнее по отношению к вирусу пространство и за пределы защитной вирусной оболочки. Микрофотография из лаборатории Майкла Б. А. Олдстоуна