Мы уже говорили о той огромной роли, которую играет эпигенетика в жизни клеток и всего нашего организма. Фактически эпигенетика занимается включением и выключением различных генов, их активацией и деактивацией. Так вот, основные механизмы эпигенетики – это, во-первых, изменение расположения гена в хромосоме, точнее его деконденсация (разупаковка) и перемещение в свободное пространство с тем, чтобы к нему могла подойти РНК-полимераза, и, во-вторых, освобождение промотора от мешающей началу работы РНК-полимеразы метильной группы (если она там была). Для особенно любознательных заметим, что существует еще несколько механизмов активации и дезактивации (экспрессии и репрессии) генов. Например, ацетилирование гистонов[45] ведет к ослаблению химических связей гистона и ДНК, стимулирует деконденсацию и активацию соответствующего гена.
В хромосомах всех живых существ выделяются две структуры, играющие важную роль в их жизнедеятельности. В центре хромосом расположены центромеры, а по ее концам – теломеры (в переводе с древнегреческого – «концевая часть»). Центромеры и теломеры представляют собой многократно повторяющиеся небольшие последовательности нуклеотидов.
Центромеры в буквальном и фигуральном смысле занимают центральное место в делении клетки. Теломеры образуют своеобразные колпачки на концах хромосом, которые защищают их от повреждений. Связанные с теломерами белки прикрепляют концы хромосомы к ядерной оболочке и иногда к ядрышкам (о них мы расскажем чуть позже). Теломерные повторы всех позвоночных состоят из шести нуклеотидов TTAGGG, повторы всех насекомых – TTAGG, повторы большинства растений – TTTAGGG.
В молодости длина теломер у человека составляет около 15 тыс. пар нуклеотидов. При каждом делении клетки длина теломер сокращается. Это объясняется тем, что ДНК-полимераза, копирующая ДНК при делении, первоначально занимает часть теломеры, к которой первоначально прикрепляется. Поэтому она не может скопировать эту часть. Клетки перестают делиться при длине теломеры 2 тыс. пар нуклеотидов, когда ДНК-полимеразе уже не на чем первоначально закрепиться. Однако человек обычно умирает раньше, с длиной теломер 5–7 тыс. пар нуклеотидов. Тем не менее связь длины теломер со временем жизни очевидна и теломерная теория старения человека остаётся одной из наиболее популярных.
В ДНК есть ещё один тип фрагментов, вызывающих у геронтологов повышенный интерес. Это транспозоны, или «прыгающие гены», – участки ДНК, способные менять свое положение в молекуле (транспозицию). У человека транспозоны составляют до 45 % всей ДНК и по типу являются ретротранспозонами. Отличительной чертой ретротранспозонов является транскрипция их кодов в РНК, как у обычных генов, а затем обратная транскрипция из РНК в ДНК в другом месте молекулы ДНК. Излишняя подвижность ретротранспозонов, их способность производить свои копии, которые могут портить ДНК, вынудила клетку использовать механизмы избавления от них. В частности, для этого используется особый тип коротких РНК – пивиРНК, помогающие специальному белку распознать опасность и дезактивировать транспозон.
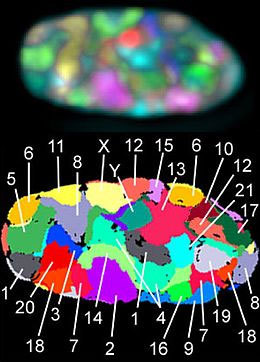
Рис. 1.2.10. Хромосомные территории (вид с использованием микроскопа – сверху и схема – внизу)
«Прыгающие гены» считаются причиной около 100 различных заболеваний. Их активность усиливается с возрастом, что внушает подозрения в том, что они могут быть важными факторами старения. Основанную на этих подозрениях теорию старения мы рассмотрим в следующей части.
Итак, ДНК в хромосоме всегда находится в более или менее компактной форме. Однако плотность укладки или степень конденсации ДНК неодинакова в различные периоды жизни клетки. Перед делением клетки конденсация ДНК, то есть плотность упаковки, значительно увеличивается. Клетка собирается перед самым ответственным моментом в своей жизни. В этот момент ДНК в хромосомах становится таким плотным, что их можно увидеть в обычный световой микроскоп. В период между делениями (в интерфазе) каждая хромосома занимает в ядре вполне определенную область (хромосомную территорию).
На рис. 1.2.10 видно (верхнее изображение), что хромосомные территории имеют нечеткие границы. Они имеют пористую границу, через которые проникают различные белки и матричная РНК. Отцовская и материнская хромосомы часто располагаются на отдаленных друг от друга территориях. Между хромосомными территориями расположены межхромосомные пространства, в которых располагаются группы ферментов, и активно идет работа генов: производится матричная РНК. Работающие гены находятся обычно в распутанном, деконденсированном состоянии и располагаются ближе к границам хромосомной территории.
В заключении этого раздела упомянем самую маленькую, но очень важную ядерную структуру – ядрышко. Оно не имеет мембраны и представляет собой сгусток белков (на 60 %) и нуклеотидов. В ДНК имеются гены, ответственные за образование специфической РНК для производства рибосом – рибосомных РНК или рРНК. Эти гены называют ядрышковыми организаторами[46]. Вблизи них располагаются ядрышки. Их число может меняться в зависимости от типа клетки, но обычно их довольно много (сотни). Основная функция ядрышек – производство рибосом, важнейших органелл, синтезирующих белки.
Как и всё в клетке, ядрышки – динамичные структуры. При начале деления клеток и конденсации (упаковке, уплотнении) хромосом они исчезают, поскольку работа генов прекращается. После деления они возникают, растут, могут сливаться друг с другом. Они могут перемещаться в межхромосомное пространство, структура которого, так же как и границы хромосомных территорий, весьма подвижна и динамична.
1.2.4. Как поддерживаются форма и структура клетки, ее энергообеспечение и управление внутриклеточными процессами
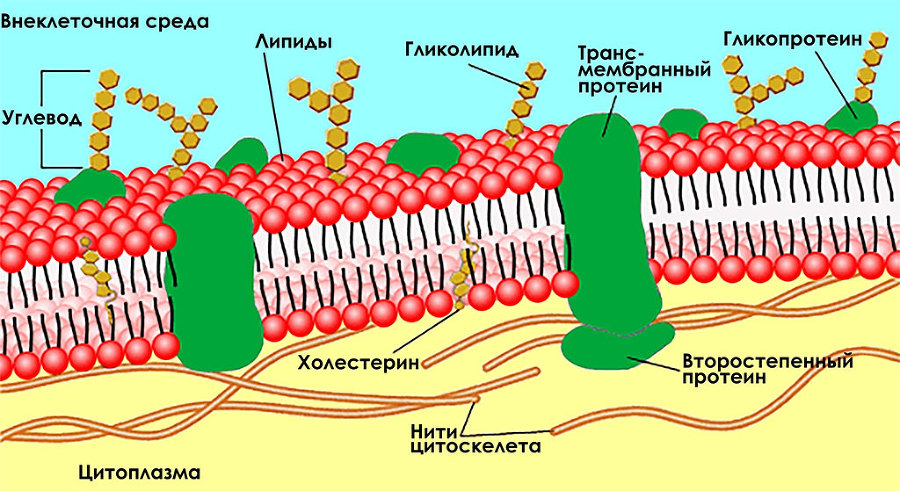
Как мы уже говорили, клетка напоминает медузу с гибкой оболочкой, наполненной цитоплазмой. Оболочка клетки – чрезвычайно сложная структура, состоящая из двух слоев жиров (липидов), погруженных в них белков, а также расположенных с внешней стороны клетки рецепторов.
Рис. 1.2.11. Схема оболочки клетки
Как видно на рисунке 1.2.11, липиды обращены гидрофильными[47] (хорошо связывающимися с водой) головками к внеклеточной и внутриклеточной среде, а гидрофобными хвостиками – друг к другу. Эти два слоя называют мембраной. Толщина клеточной мембраны – 5–7 нм. Мембрана – универсальная структура. Такая же мембрана ограждает ядро и различные органеллы. При этом неверно представлять себе мембрану как стену из липидов. На самом деле это чрезвычайно подвижная и гибкая оболочка с множеством встроенных между липидами белков, образующих различные проходы и поры. Относительно небольшие молекулы – глюкоза, аминокислоты, жирные кислоты и ионы – могут проникать через мембрану. Проникновение идет самопроизвольно от областей с большей концентрацией к областям с меньшей концентрацией. Однако эти процессы могут контролироваться мембраной.
Другие молекулы могут проходить только при помощи трансмембранных белков, образующих контролируемые проходы в оболочке. Обычно прохождение таких молекул требует затрат энергии. Примером прохождения веществ от областей с меньшей концентрацией к областям с большей концентрацией, требующим потребления энергии, может служить калиево-натриевый насос, представляющий собой фермент Na+/K+-АТФ-аза. Этот фермент присоединяет с внутренней стороны мембраны три иона Na+. При помощи получаемой извне от молекулы АТФ энергии три иона Na+ переносятся на внешнюю сторону мембраны, где они отщепляются и присоединяется два иона К+. Один заряд лишний. Так возникает разность потенциалов между внешней и внутренней частью оболочки клетки. После этого фермент возвращается в исходную позицию, а ионы К+ оказываются на внутренней стороне мембраны. В результате постоянного действия калиево-натриевого насоса концентрация калия в наших клетках в 30 раз больше, чем в плазме крови, а концентрация натрия, наоборот, в 15 раз меньше, чем вовне. Na связывает воду, а K пытается вывести ее из клетки. Этот насос обеспечивает циркуляцию жидкости из межклеточного пространства в клетку и обратно. Вместе с ней циркулируют и питательные вещества внутрь клетки, а из клетки – продукты жизнедеятельности клетки. Работа калиево-натриевого насоса потребляет примерно треть всей энергии, расходуемой клеткой.