Причины замедления роста приспособленности связаны с тем, что каждая закрепившаяся полезная мутация делает последующие генетические усовершенствования в среднем менее полезными. Авторы называют этот эффект “эпистазом убывающей доходности” (по аналогии с экономическим законом убывающей доходности). Иными словами, сначала происходят наиболее радикальные адаптивные изменения, а затем идет все более тонкая настройка и оптимизация фенотипа.
Еще один фактор, способствующий замедлению роста приспособленности и тесно связанный с предыдущим, – клональная интерференция, то есть конкуренция между клонами бактерий с разными полезными мутациями. Напомним, что бактерии, участвующие в эксперименте, – бесполые, они не могут обмениваться генами. Поэтому, если у одной бактерии возникнет одна полезная мутация, а у другой – другая, эти мутации не смогут объединиться в одном геноме (как это произошло бы у нормальных микробов, способных к горизонтальному переносу генов, и уж тем более у организмов, размножающихся половым путем; см. Исследование № 7). Вместо этого потомки первой бактерии начнут не на жизнь, а на смерть (не на свою, конечно, а своей линии) конкурировать с потомками второй. Победит, естественно, тот клон, чья мутация окажется полезнее. Вторая, менее полезная мутация будет вытеснена и потеряна. Придется теперь ждать, пока она случайно появится снова у бактерии, уже имеющей первую мутацию. А закрепиться она сможет, только если у нее не окажется более удачливых конкурентов. Клональная интерференция во многом объясняет, почему на первых этапах эксперимента фиксировались в основном мутации с сильным полезным эффектом (в среднем первая закрепившаяся в каждой популяции мутация повышала приспособленность на 10 % – это очень много), а затем уже постепенно начинали фиксироваться все менее и менее полезные мутации.
Разумеется, у бактерий, способных к горизонтальному переносу генов, адаптация шла бы гораздо быстрее (см. Исследование № 7). Но и анализировать результаты было бы труднее, потому что к мутационной изменчивости добавилась бы комбинативная, а на эволюцию стали бы влиять такие факторы, как наследственные различия по склонности к приему и передаче генов, избирательность при выборе партнеров и т. д.
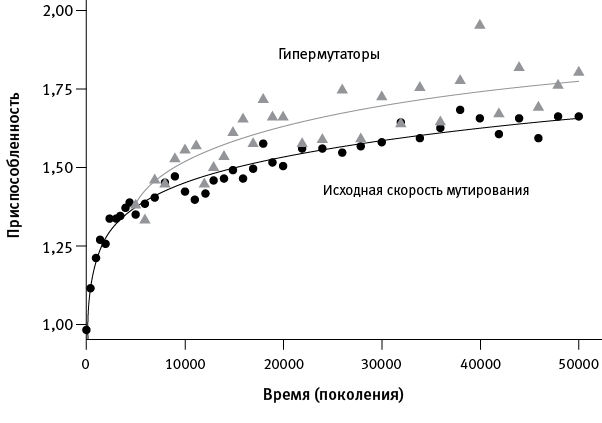
Еще один важный результат дало сравнение роста приспособленности у популяций, сохранивших исходный (низкий) темп мутагенеза, и тех, где закрепились аллели-мутаторы, несущие мутации, которые резко (в среднем на два порядка) повысили темп мутирования. На сегодняшний день “гипермутаторами” стали шесть из двенадцати популяций.
рис. 1.2. Повышение темпа мутагенеза ускоряет адаптацию. Черные точки – усредненные данные по шести популяциям, в которых темп мутагенеза остался на исходном низком уровне. Серые треугольники – усредненные данные по трем популяциям, в которых на ранних этапах эксперимента (за первые 20 000 поколений) закрепились аллели-мутаторы. Видно, что приспособленность у вторых росла быстрее, чем у первых. Еще три популяции, у которых мутаторы закрепились поздно, в данном случае не рассматривались. По рисунку из Wiser et al., 2013.
Оказалось, что у “гипермутаторов” адаптация протекала ускоренными темпами (рис. 1.2). Получается, повышенный темп мутагенеза пошел бактериям на пользу. Это противоречит распространенной идее о том, что в стабильных условиях организмам выгодно снизить темп мутирования до нуля. Ведь большинство вновь возникающих мутаций вредны, поэтому мутация, повышающая темп мутагенеза, в краткосрочной перспективе приносит больше вреда, чем пользы, и отбор, по идее, должен действовать против нее.
Почему же аллели-мутаторы все-таки распространяются? Дело в том, что в бесполой популяции они часто спасаются от отбраковки благодаря эффекту, который называют генетическим автостопом. Если какой-то гипермутабельной бактерии повезет и у нее появится редкая и очень полезная мутация, отбор начнет поддерживать последнюю. При этом вместе с полезной мутацией будет автоматически распространяться и сидящий в том же геноме аллель-мутатор. Он будет, подобно пассажиру-безбилетнику, ехать прицепом к хромосоме, распространение которой в генофонде популяции “оплачено” полезной мутацией. А все потому, что в бесполой популяции, где нет перетасовки генов и хромосомных участков между отдельными клетками, отбираться могут только целые геномы, но не отдельные гены.
В популяции, способной к генетическому обмену, ситуация будет иной. Связка мутатора с полезной мутацией не будет неразрывной, что позволит отбору работать с ними индивидуально. В итоге отбор сможет закрепить полезную мутацию и отбраковать породивший ее аллель-мутатор (см. Исследование № 7). Но это пока лишь теория: эволюционный эксперимент на организмах, способных к генетическому обмену, сопоставимый по масштабу с экспериментом Ленски, еще не поставлен.
Итак, мы увидели, что предела приспособленности, по всей видимости, нет. Об этом нам говорят экспериментальные данные. Этот вывод кажется контринтуитивным, потому что “высшую точку” приспособленности можно легко себе представить как нечто реальное и достижимое. Казалось бы, для заданных постоянных условий должно существовать некое оптимальное, наиболее приспособленное состояние – такое, что у организма, его достигшего, никакая мутация уже не сможет повысить приспособленность. Даже если для реальной эволюционирующей популяции этот оптимум недостижим, он продолжает оставаться для биологов удобной абстракцией, упрощением, помогающим решать определенные задачи. Эволюционисты привыкли представлять себе этот оптимум в виде горной вершины на воображаемом ландшафте. Но при этом важно понимать, какие пути ведут к высшей точке, или, образно выражаясь, какой рельеф имеют склоны горы. Упрощенное понимание естественного отбора рисует нам гладкие склоны и прямой путь к вершине. Но это, очевидно, не соответствует биологическим реалиям. Благодаря стремительному развитию науки, “путь к вершине приспособленности” постепенно перестает быть абстракцией и начинает поддаваться картированию. Об этом – следующая глава.
Исследование № 2
Трудный путь к совершенству по ландшафту приспособленности
“Ландшафт приспособленности” – это воображаемый график, показывающий, как зависит функциональность гена от его нуклеотидной последовательности (или работоспособность белка от его аминокислотной последовательности). Биологи привыкли считать ландшафт приспособленности абстракцией, полезной для теоретических рассуждений, но недосягаемой для реального изучения. Однако стремительное развитие биотехнологий уже позволяет картировать небольшие области ландшафта приспособленности конкретных белков. Американские биологи экспериментально изучили свойства всех возможных аминокислотных комбинаций, занимающих четыре ключевые позиции в одном из белков кишечной палочки. Неожиданно оказалось, что из 160 000 комбинаций работоспособны целых 1659 (более 1 %). При этом эволюционные маршруты от одних “разрешенных” последовательностей к другим, как правило, оказываются длинными и окольными. Это связано с сильным эпистазом – зависимостью пользы мутации от того, какие мутации успели закрепиться раньше. Иными словами, важно не только само появление тех или иных мутаций, но и порядок их появления. Возможно, из-за эпистаза эволюция не может найти многие удачные решения, а ее пути становятся до некоторой степени предсказуемыми.


Одним из способов представить себе, как идет эволюция, является картирование ландшафта приспособленности конкретных белков. В схематичном виде ландшафт приспособленности показан на илл. i (см. цветную вклейку). Конечно, в идеале хотелось бы просканировать все “пространство последовательностей” (все возможные аминокислотные последовательности белка), определив для каждой его точки (то есть для каждой последовательности) эффективность выполнения белком интересующей нас функции. Однако такая задача невыполнима, поскольку даже для маленькой белковой молекулы в 100 аминокислот число точек в пространстве последовательностей (то есть число всех возможных белков такого размера) на много порядков превышает число атомов во Вселенной (первое равно 20100 ≈ 10130, второе – в 1050 раз меньше).