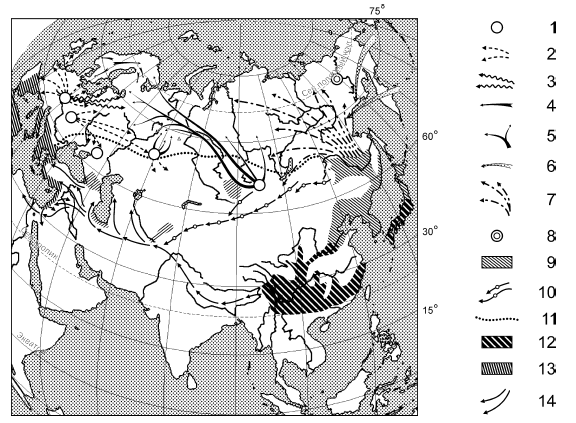
Рис. 8А. Схема расположения центров и путей распространения флор (по: Алёхин, Кудряшев, Говорухин, 1961) в Евразии:
Европейско-Сибирские лесные флоры: 1 – ледниковые убежища жизни и центры распространения флоры широколиственных и смешанных лесов и таёжной флоры; 2 – послеледниковые пути расселения флоры широколиственных лесов; 3 – пути расселения флоры хвойно-широколиственных лесов; 4 – скандинавские послеледниковые пути расселения флор еловых и сосновых лесов; 5 – пути послеледникового расселения таёжной флоры из Алтае-Саянского и Южно-Уральского убежищ жизни; 6 – расселение лесной флоры Берингии; 7 – пути расселения флоры светлохвойных (б.ч. лиственничных) и смешанных лесов Восточной Сибири из Северного Китая в третичном и четвертичном периодах; 8 – древний третичный центр происхождения флоры сибирских и американских хвойных лесов; Восточно-Азиатские и Средиземноморские лесные флоры: 9 – области реликтовых флор (третичных) листопадных и смешанных лесов Азии; 10 – плейстоценовые пути расселения флоры азиатских широколиственных лесов (Алтае-Саянский путь); 11 – северные (сибирские и урало-украинские) пути расселения третичных флор широколиственных лесов; 12 – флоры третичных субтропических вечнозелёных лесов Китайско-Японской подобласти и их реликты на Кавказе; 13 – флора Западной Средиземноморской подобласти; 14 – пути расселения флоры вечнозелёных и листопадных лесов из Юга Восточной Азии в Средиземноморскую подобласть в третичном периоде (Гималайский путь).
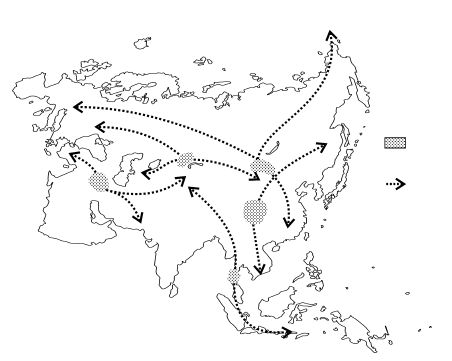
Рис. 8Б. Схема расположения центров и путей распространения фаун (по: Мекаев, 1987) в Евразии:
1 – ледниковые рефугиумы флоры широколиственных, смешанных и таёжных лесов; 2 – пути расселения флоры широколиственных лесов в постледниковье; 3 – пути расселения флоры хвойно-широколиственных лесов; 4 – пути расселения флоры хвойных лесов; 5 – области реликтовых флор третичного времени; 6 – фаунистические центры; 7 – пути расселения фаун.
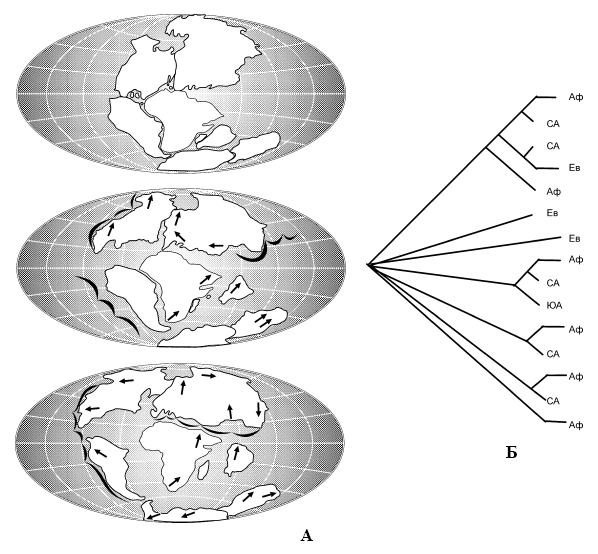
Третье направление в биогеографии – теория викарианса (мобилизма). Она имеет в основе постулат, который гласит, что схемы филогенетического (эволюционного) порядка замещения таксонов и исторической последовательности трансформации структуры географической арены строго согласованы. Наиболее ярким примером мобилистической биогеографии является согласование эволюционных схем развития жизни с рассредоточением фрагментов мезозойского суперконтинента Пангея в ходе геологической истории планеты, рисуемое геофизиками (Еськов, 1984; Кафанов, Кудряшов, 2000). В соответствии с канонами обсуждаемой теории, биологические таксоны сначала расставляются с помощью кладистического анализа по степени отклонения от исходной формы. Затем по аналогии с кладограммами систематиков строятся биогеографические кладограммы, показывающие не только современную картину распространения какого-либо таксона, но и историческую последовательность её формирования (рис. 9).
Рис. 9.
А – схема изменений местоположения континентов (по: А. Вегенеру из: Клечек, Якеш, 1985); Б – географические кладограммы лошадей по современным и ископаемым видам (по: Биосфера, 1988):
Аф – Африка, Ев – Евразия, СА – Северная Америка, ЮА – Южная Америка.
В качестве краеугольных камней викариантной (мобилистической или кладистической) теории рассматриваются следующие постулаты:
1) если таксон имеет ареал, разорванный какой-нибудь преградой, то его предки расселялись тогда, когда этой преграды ещё не было;
2) степень родства двух ныне изолированных контингентов таксона соответствует времени их изоляции, т. е. наиболее близкородственные из них представлены на наиболее поздно разошедшихся фрагментах некогда единого ареала;
3) более примитивные представители таксона всегда располагаются географически ближе к первичному центру происхождения, чем более продвинутые (Красилов, 1977).
Согласно императиву мобилистической (викариантной) биогеографии, современная биогеографическая картина составляет только преходящий эпизод в перманентной череде сочетаний биологических видов, замещающих друг друга в ходе эволюции, адекватно порядку изменения структуры географической арены. Благодаря неодинаковой скорости изменений в разных её частях единовременно, но разобщённо в пространстве сосуществуют серии заместивших друг друга таксонов, что обеспечивает гетерогенность флор и фаун. В какой-то мере это явление напоминает смену актёров на одну и ту же роль в киносериале по ходу изменения конъюктуры актерского рынка в разные периоды съёмок. Зритель же вынужден мириться со сменой лиц персонажей от серии к серии, утешаясь неизменностью сюжетной линии (рис. 9).
Четвёртое направление биогеографии опирается на теорию динамического равновесия, сконструированную американскими учёными Мак-Артуром и Уилсоном (1967) для нужд биогеографии островов. Суть теории заключается в том, что количество видов, населяющих остров, определяется равновесием между иммиграцией и вымиранием. Сколько видов вымирает – столько замещается за счёт вселения. В конечном итоге общее число видов на острове определённой площади и удаления от очага иммиграции остаётся приблизительно одинаковым (рис. 10). Если острова разной площади расположены на разном расстоянии от источника колонизации, то увеличение расстояния уменьшает скорость иммиграции, а увеличение площади острова – уменьшает скорость вымирания. Зависящее от площади острова равновесное число видов изменяется при этом по степенному закону. Поскольку островной эффект, в принципе, имеет место не только в океане, но и на суше (озёра как острова воды среди сухопутья, пятна степей среди тайги), то применение теории равновесия выходит за рамки морской биогеографии, её породившей.
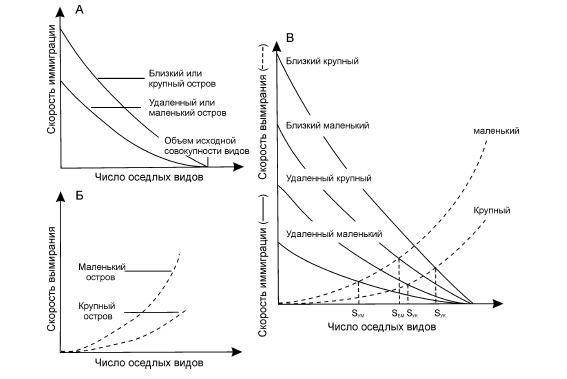
Рис. 10. Теория равновесия Мак-Артура и Уилсона (по: McArthur, Wilson, 1967 из: Бигон и др., 1989):
А – зависимость скорости иммиграции от числа обосновавшихся видов для крупных и мелких островов, лежащих вблизи и вдали от континента; Б – зависимость скорости вымирания от числа обосновавшихся на острове видов для крупных и мелких островов; В – баланс между вымиранием для крупных и мелких и для близких и удалённых от континента островов. В каждом случае S – обозначает равновесное видовое богатство.
Императивом экологической биогеографии становится уверенность, что замещение таксонов, как эволюционное изменение форм путём естественного отбора, так и комбинаторное заимствование подходящих форм в пространстве из имеющегося по соседству ассортимента, происходит в любом месте географической арены обязательно под контролем конкретных экосистем, где все элементы скованы, как каторжники, единой цепью круговорота веществ и энергии. Даже при появлении новой, замечательной по своим потенциям формы жизни, её внедрение в какое-либо местообитание возможно лишь с “разрешения” экосистемы при наличии подходящей свободной экологической ниши.
1.5. Методы биогеографии
Несмотря на отмеченную специфику, все отрасли биогеографии роднит главное: все они изучают живые системы любого уровня организации с единых хорологических позиций, с помощью сравнительно-географического метода на базе количественных и качественных учётов встречаемости организмов в прошлом или настоящем.
Являясь наукой пограничной, биогеография опирается на главный метод и широко пользуется результатами исследований соседних отраслей знания. Уже было сказано о значении кладистического анализа, заимствованного из биосистематики. Находят применение кариологические методы. Широко используется изучение макроскопических остатков и микрофоссилий (игл, раковин фораминифер, спор, пыльцы растений). Переносящаяся в атмосфере пыльца может оседать на поверхности субстратов, благоприятных для её фоссилизации, при быстром погребении в условиях, способствующих её сохранению (на торфяниках, днищах озёр, в некоторых почвах). Определение и подсчёт зёрен пыльцы в образце, взятом через много лет, даёт возможность воссоздать средний состав и направление пыльцевого “дождя” в ту или иную эпоху, представить себе палеопейзаж и сравнить его с современным.