Рис. 4.2. Влияние возраста на растворимость коллагена сухожилия хвоста (промытого фосфорной кислотой) в воде при 65 °C в течение 10 мин для интактных крыс (I) и крыс, лишенных гипофиза (II) [20]
Рис. 4.3. Увеличение с возрастом суммарного веса коллагена в сухожилии хвоста и количества нерастворимого коллагена [16]. С помощью гель-фильтрации образцов на биогеле А-15 показано увеличение с возрастом количества нерастворимого коллагена и соответствующее уменьшение α-субъединиц
Рис. 4.4. Распределение α-цепей и коллагена других типов в коже крыс разного возраста [36]
Рис. 4.5. Зависимость веса груза, который может поднять сухожилие крысы при сокращении, от возраста [89]. Предел прочности при растяжении (или сила сокращения) равномерно увеличивается с возрастом
Рис. 4.6. Увеличение с возрастом времени, которое требуется для разрыва изолированного волокна коллагена из сухожилия хвоста крысы в 7 М мочевине при 40 °C и рН 7,0; вес груза 2 г [22]
Коллаген является главным органическим компонентом экстрацеллюлярного матрикса, в особенности соединительных тканей, таких, как кости и хрящи. Он служит как бы "лесами" для клеток при построении этих тканей и выполняет механические и защитные функции.
При старении млекопитающих обычно наблюдаются увеличение числа морщин на коже, уменьшение костной массы и увеличение хрупкости костей, а также изменение физических свойств хрусталика. Происходит ли это благодаря появлению с возрастом новых типов коллагена или благодаря прогрессирующим посттрансляционным изменениям в макромолекулах коллагена, вызывающим потерю функций ткани? Если верна первое, то это может означать, что перечисленные выше изменения свойств коллагена происходят из-за изменений первичных центров, т. е. в пожилом возрасте происходит экспрессия генов, ответственных за синтез нерастворимого коллагена. Если же верно второе предположение, то причина старения имеет вторичный характер. Изменения физических свойств коллагена по мере старения животного в этом случае могут происходить из-за изменений с возрастом уровней эффекторов, что приводит к модификации свойств коллагена уже после его синтеза. Таким образом, для того чтобы разобраться в изменении свойств коллагена как функции возраста, необходимо знать его молекулярную структуру.
Молекулярная структура
Внеклеточный коллаген представляет собой фибриллы, состоящие из нескольких мономеров. Каждый мономер — это тройная спираль из трех полипептидных цепей, свернутых друг относительно друга. В коллагене разных тканей найдено два типа полипептидных цепей — α1 и α2. Цепи α1 имеют четыре варианта — α1(I), α1(II), α1(III) и α1(IV). Четыре варианта α1 и цепи типа α2 различают последовательностью аминокислот и, следовательно, кодируются разными генами.
Каждая α-цепь состоит из ~1050 аминокислотных остатков и имеет мол. массу 100000. Приблизительно 15–25 остатков в каждом из NH2- и COOH-концевых участков α-цепи имеют последовательность, не свойственную коллагену и не имеющую формы спирали. Эти концевые участки называются: телопептидами. В отличие от центральных спиральных областей цепи, характерных для коллагена, они не содержат Gly в каждом третьем положении. На NH2-конце находится пептид Lys9, сохранившийся в ходе эволюции. Он превращается вне клетки в аллизин, принимает участие в образовании поперечных сшивок с α-цепью другого мономера коллагена и, следовательно, в формировании фибриллы. COOH-концевая область также содержит сохранившийся в ходе эволюции Lys1044, участвующий в образовании сшивки внутри мономера и в формировании фибриллы [57].
Центральная часть α-цепи состоит из ~1000 остатков, соединенных в повторяющейся последовательности Gly — X-Y, т. е. Gly занимает каждое третье положение. Таким образом, всего в α-цепи около 337 глициновых остатков, составляющих одну треть всех аминокислот коллагена. Положение X и Y занимают другие аминокислоты. Часто встречаются также аминокислоты пролин (Pro) и оксипролин (Hyp). Pro занимает положение X, а Hyp — положение Y. Обычно Hyp гидроксилирован по четвертому углеродному атому. Разные типы α-цепей отличаются друг от друга аминокислотами в положениях X и Y. Наличие Gly и его присутствие в каждом третьем положении характерны для всех цепей, тогда как содержание Hyp и Pro в разных α-цепях различно.
Чем выше суммарное содержание Hyp+Pro, тем выше температура плавления (Tm) коллагена и его стабильность (Tm — температура, при которой разрушается 50 % спиральной структуры):
В отличие от глобулярных белков α-цепи не имеют водородных связей внутри цепи. Таким образом, в них отсутствуют α-cпирали и они представляют собой β-структуры. Если у глобулярных белков, имеющих α-спиральную структуру, приходится 5,4 аминокислотных остатков на виток, а подъем на один остаток равен 0,15 нм, для α-цепей коллагена эти значения составляют 3,3 остатка и 0,29 нм соответственно. Каждая цепь имеет левые витки с шагом 0,9 нм, содержащим три остатка.
Три α-цепи, уложенные параллельно, образуют тройную спираль мономера коллагена. Вокруг центральной оси они расположены таким образом, что каждая закручена относительно другой влево. Три цепи удерживаются друг возле друга водородными связями. Несмотря на то что энергия Н-связей составляет всего 3–5 ккал/моль, существование большого числа таких связей (две Н-связи на три остатка) обеспечивает удерживание трех цепей рядом, причем именно водородные связи ответственны за структуру тройной спирали. Это оказывается возможным благодаря наличию в каждом третьем положении Gly. У глицина нет боковых групп, а его атом водорода при α-углеродном атоме расположен в пространстве между цепями. Азот иминогруппы Gly одной цепи связан Н-связью с карбонильным (кислородом Gly другой цепи (рис. 4.7). Любая другая аминокислота препятствовала бы образованию Н-связи между цепями и закручиванию цепей. Hyp также образует Н-связи между цепями через молекулы воды и, следовательно, тоже вносит вклад в стабильность мономера [63]. Коллаген, в котором Pro не гидроксилирован, имеет Tm в интервале 20–25 °C. Если Pro гидроксилирован, то Tm коллагена равна 37 °C. Таким образом, структура тройной спирали коллагенового мономера поддерживается в значительной степени благодаря водородным связям.
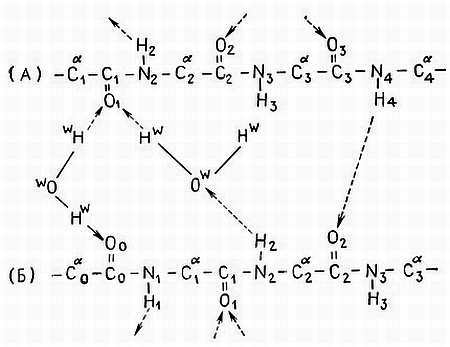
Рис. 4.7. Водородные связи в тройной спирали коллагена [63]. Предполагается образование водородных связей двух типов: 1) однократно связанных структур между группами NH цепи А и С=О цепи Б; 2) дважды связанных структур с водным мостиком между группой NH цепи Б, связанной водородной связью с кислородом молекулы воды, которая в свою очередь связана с кислородом цепи А. Кроме того, вторая молекула воды связывает атомы кислорода цепи А и цепи Б